In this article we will discuss about:- 1. Echolocation in Bats 2. Sound Production in Bats 3. Characteristics of Echolocation Sounds 4. Working Space and Information Content of Echoes.
Echolocation in Bats:
Echolocation is the emission of high frequency sound (ultrasonic sound, about 20 kilohertz) which is utilised for detecting the presence of objects (including food) by the echoes produced. It is a good substitute for vision for those animals, such as bats, which have to hunt in darkness. These high frequency sounds are produced in the larynx (voice box) and are generally restricted in insectivorous bats of the suborder Microchiroptera.
The mega-chiropterans, that feed on fruit, flower, nectar etc., generally lack ultrasonic orientation. However, some of its members (Rousettus and, perhaps, Epomophorus wahlbergi) produce orientation sounds that are largely audible and have ultrasonic components that are produced not in the larynx but by clicking the tongue. Bats are not the only mammals possessing ultrasonic orientation sounds. However, in case of bats, it has reached its peak.
All bats, however, possess and utilise low frequency sounds (below 20 kHz — within normal audible range of humans), which are generally used to facilitate social interactions such as territorial spacing among individuals, mother-infant communication, recognition and warning calls.
ADVERTISEMENTS:
The echolocation mechanism adopted is not fully understood and certainly is not the same. Lazzaro Spallanzani (1729-1799), an Italian at the age of 64, first experimented and proved that bats did not rely upon their visual sense to fly and capture prey at night. He, however, could not provide a plausible explanation.
In 1920, H. Hartridge reconfirmed the notion of Spallanzani that bats oriented with auditory rather than visual cues. Later, in 1958, Donald Griffin, a Harvard biology student, showed conclusively that the bats emit pulses of high- frequency sounds which strike objects and reflect back to the animals. Thus, bats hunt by echolocation and not by listening to the sounds made by the insects.
Anatomical Structure in Sound Production and Reception:
A. Sound Production:
Micro-chiropterans generate ultrasonic sounds in the larynx. The larynx in micro-chiropterans is proportionately longer than that found in mega-chiropterans and the cartilages are ossified to make a rigid framework. The strong cricothyroid muscles put greater tension on the light vocal cords. The complexity and large size of this sound-producing structure directly relates to the range of ultrasonic sound that it produces.
It is performed by discrete pulses of high intensity and up to 150 kHz frequency which is emitted through the nose and mouth. In the case of horseshoe bats (Fig. 5.19A) there are special resonating chambers and the face is elaborately modified forming a nose-leaf.

B. Role of Nose-leaf:
The role of nose- leaf in echolocation is not well-understood. Rhinolophid and hipposiderid bats possess complex nose-leaf that serve to beam the ultrasonic sounds and help in directing the signal to a particular direction. The complex flaps and folds also serves to shield the ears from the outbound, nasally emitted signals and so increase their overall sensitivity to returning echoes.
C. Specialisation of Ears:
The ears of bats are greatly specialised (Fig. 5.19). The pinnae or external ear of most echo-locating bats are large and funnel-shaped. They are small in bats that fly fast and emit loud sounds, while they are enormous in those that hunt insects using faint pulses.
The ears of many bats contain a short to long vertical flap (tragus) that may improve the directionality or sensitivity to inbound echoes. If may also act as a deflective shield that protects the ear from the intense outbound acoustic pulses.
ADVERTISEMENTS:
The pinnae are positioned in such a way so that it can be receptive to inbound echoes from 30°-40° on either side of the mouth. The direction of an obstacle or target is enhanced by comparing the differences in the intensity and quality of echoes as they arrive at each ear.
As there are two ears, it yields stereophonic images in a manner like that of our two eyes which yield stereo-visual images. Rhinolophus is able to move the ears back and forth alternately when it scans its surroundings.
The middle ear comprises of the tympanic bone (ring) that supports the tympanic membrane and the middle ear ossicles (Fig. 5.20). The thickness of the tympanic membrane of micro-chiropterans ranges from 0-002 to 0-011 mm, which is relatively thinner than the eardrums of other mammals.
A correlation seems to exist between frequency sensitivity in hearing and the acoustic signals emitted during echolocation. Bats that operate with high frequencies (50-125 kHz) generally have a smaller tympanic area than bats that operate at lower frequencies (below 50 kHz). Similar correlation exists with the mass of the middle ear ossicles.

The middle ear has two muscles—tensor tympani and stapedius, which are much developed in micro-chiroptera (Fig. 5.20B). Of the two, the stapedius muscle is especially important in regulating the sound entering the ears.
In the Mexican Free-tailed bat (Tadarida brasiliensis), the stapedius muscle begins to contract about 10 ms before the emission of pulse. This causes the stapes bone to be pulled away from the oval window which momentarily deafens the bat to its own pulse emission. The stapedius muscle relaxes after the pulse is emitted and the sensitivity is restored. The ear, thus, is ready to receive an echo.
The cochlea and associated labyrinths are the most important regions of the ear. In micro-chiroptera, the cochlea is relatively larger and more complex in structure (Fig. 5.20A-D). In most micro-chiropterans, the number of turns in the cochlear duct varies between 2-5 and 3, while in Rhinolophus it is 3-5.
The membranous labyrinth and organ of corti (Fig. 5.20C and D) are specialised anatomically and physiologically for the reception of certain frequencies. The cochlea, due to these adaptive features, are somewhat conical in shape instead of spherical. To reduce the potential interference from outbound pulses, the cochlea is suspended loosely in connective tissue, rather than being solidly fused to the bottom of the skull.
D. Specialisation of Brain:
ADVERTISEMENTS:
The cerebral hemispheres are small and the olfactory portions reduced, but the inferior collicula (concerned with hearing) and cerebellum are large (Fig. 5.20E). The eyes, moderately large and the retina containing mainly rods are used probably during twilight.
Electrical nerve impulses from the sensory cells in the organ of Corti are collected and travel along the auditory nerves to the inferior colliculi, where the sounds are analysed for its information content. The location, large size and general exposure of the inferior colliculi on top of the midbrain has led to its hearing range extending from a frequency of less than 1 kHz to about 200 kHz.
The cells of the colliculi responds in the following manner:
(i) Some cells of the colliculi responds weakly to outbound signals, but responds strongly to the returning echo.
(ii) Certain cells respond to specific frequencies in the echo.
(iii) Other cells are particularly sensitive to the time delay between the outgoing pulse and the returning echo.
E. Ontogenetic Development of Echolocation:
In many species of micro-chiropterans the nursery roost has a crowded condition. This means that the reunion between a returning mother and infant is often difficult and must occur amidst a cacophony of other mother and infant calls.
Infant bats produce many locational or isolating signals, which are characteristically lower in frequency than that of the adults. These are repeated on a regular basis and often consist of one long and one short modulated pulse.
For instance, the infants of mouse-eared bat, Myotis blythii (Fig. 5.19B), emit pulses lasting about 25 to 50 mls. These are repeated at intervals that may vary from 100 to 400 ms. The frequency range of these isolation calls varies from 14-16 kHz to about 22-24 kHz.
When the mother returns to the roost site, she apparently picks up these isolation signals of her infant. She in return emits a signal which triggers a vocal exchange between mother and infant.
As young bats grow older they reduce the use of isolation calls and gradually raise the frequency range of their calls. The pulse duration and pulse repetition rate also becomes much closely similar to those of adult bats.
Characteristics of Echolocation Sounds:
Before going into the novelty of chiropteran acoustic orientation, some physical parameters of sound are discussed:
A. Physical Aspects of Sound:
Sound is a form of energy which is manifested in the form of disturbance or vibration in a medium. Sound can be measured in a number of ways.
(a) Wavelength and frequency:
Sound can be measured in frequency which is expressed as Hertz or cycles per second (Fig. 5.21). Another way sound can be characterised is by its wavelength (Fig. 5.21), which is inversely related to its frequency. High frequency sound has a short wavelength. For example, a sound frequency of 50 kHz has wavelength of only 6-8 mm, while a low frequency sound of 34 Hz has a wavelength of about 10 m.
Bats utilise high frequency sounds rather than those of low frequency because:
(i) High frequency sounds get weakened in intensity as they pass through air, whereas low frequency sounds retain their intensity over long distances.
(ii) By using high frequency sounds bats avoid possible interference from environmental background noises such as insect songs, wind or other nocturnal noises that are generally in the low frequency range.
(iii) By utilizing high frequency signals, it provides a means of escaping detection by potential nocturnal predators.
(iv) It may be a way of detecting insect prey without being detected in turn.
It is to be noted that some moths who fall prey to bats, can hear bat’s sounds and exercises evasive maneuvers or even produces high frequency sounds of their own to jam an approaching bat’s course.
(b) Amplitude or intensity:
Sound contains energy-amplitude, intensity or loudness. Amplitude is represented by the height above and below the zero reference line of the wave form, as shown in Fig. 5.21. It can be estimated by measuring the difference in pressure between the peak and the trough of the wave. This pressure is expressed as Newtons per square meter (Nm−2).

(c) Harmonics or overtones:
Natural sounds are complex in structure and consist of a collection of several to many frequencies. The different frequencies in a complex sound are called harmonics or overtones. The harmonic structure of a complex sound is not a random assortment of frequencies, but it is rather an ordered sequence of related frequencies (Fig. 5.22).

The complex sound of Fig. 5.22C consists of two related frequencies. The lowest in this series (Fig. 5.22A) and thereby the fundamental frequency is 1,000 Hz. The next frequency, or second harmonic (Fig. 5.22B) is higher than 1,000 Hz and is precisely 2,000 Hz.
Complex sounds are further complicated by the fact that each harmonic may be produced with different amplitudes or intensities, and, in some cases, harmonics may be ‘dropped out’ of the sound spectrum altogether.
Harmonics of bats:
Bats produce a variety of complex sounds in their acoustic orientation vocalizations, which comprises of sweeps through several frequencies with accompanying harmonics. These sounds are produced in the larynx and may be filtered or altered by various resonating cavities or other peculiarities in the vocal tract or nasal passages. The acoustic sounds thus emitted by the bat have a particular structure.
By using orientation sounds with multiple harmonics, bats increase the discriminatory ability of the sound as each harmonic represents a discrete frequency with its own wavelength and target size recognition characteristic.
Such range of harmonics would be of definite value in hunting in a dense, tangled environment (or along solid surfaces) by improving the contrast between echoes reflected from a potential prey item and its background.
B. Echolocation Sounds Produced by Micro-Chiropterans:
Micro-chiropterans produce high frequency sounds that are acoustic signals emitted by the bat for the purpose of gathering vital information about objects in its immediate vicinity. The nature of this biological imaging system, its quality and quantity, varies widely among the various species of micro-chiropterans.
The micro-chiropterans ultrasonic signal is characterised by a combination of several parameters such as:
(i) Duration of signal:
The orientation signal used by bats varies in duration from about 0-2 to 200 milliseconds (ms). The range of this variation is divided into 1. those that are exceptionally short (less than 2 ms), 2. those that are of moderate duration (less than 10 ms but more than 2 ms), and 3. those that are very long (more than 10 ms).
(ii) Harmonic structure of signal:
Bats can incorporate from one to about 5 harmonics in their acoustic orientation signals. Contrary to the above, some species do not incorporate any harmonics in their echolocation sounds.
(iii) Frequency-modulated (FM) component of signal:
The frequency-modulated components comprise of sweeps that starts with a beginning frequency, passes through several to many intermediate frequencies and, ultimately, ends in some ending frequency that is usually lower than the beginning frequency.
FM components may be:
1. Narrow in band width that sweeps through relatively few frequencies (for example, 55 kHz to 40 kHz = bandwidth 15 kHz), or
2. Broad in bandwidth that sweeps through many frequencies (for example, 100 kHz to 50 kHz = bandwidth 50 kHz).
(iv) Constant-frequency (CF) component of signal:
Constant-frequency signals are echo- location sounds emitted at one frequency. Many bats incorporate a CF component in their acoustic orientation calls.
(v) Sequence of frequency components in signal:
Both CF and FM components are commonly encountered in ultrasonic sounds of micro-chiropteran. Any one of the components may precede the other.
(vi) Amplitude of signal:
The amplitude (intensity or loudness) of the acoustic orientation sounds of bats are of a wide range. Intensities of 0-1-0-32 Nm−2 have been recorded for the Greater False vampire (Megadermalyra). While the Greater Horseshoe bat and the Mediterranean Horseshoe bat produces intensities of 27-0 Nm−2 and 2.0 Nm−2, respectively. The Little Brown bat (Myotis lucifugus) produces sounds between 6-0 and 17-3 Nm−2.
(vii) Variations of repertoire:
Each bat may change its acoustic repertoire along one or more of the parameters discussed above. This change may be continuous in nature or it may differ in different foraging situations.
Micro-chiropterans acoustic imaging system is of two general kinds:
1. Time frequency spectrum consisting primarily of broadband, frequency-modulated (FM) signals.
2. Time-frequency structure that consists mainly of narrowband, constant frequency (CF) signals.
(a) Broadband, frequency-modulated signals:
The first broadband acoustic pattern consists of frequency-modulated signals with multiple harmonics. The acoustic orientation of phyllostomids (New World leaf-nosed bats) has been well-studied and they are the most diverse with regards to their feeding habits. Their ultrasonic signals are exceptionally low in amplitude (less than 0-1 Nm−2) and of short duration (0-5-2-5 ms).
Although the band width of each harmonic is relatively narrow (average 20-30 kHz), the abrupt linearity of the frequency sweep causes these harmonics to overlap, thereby creating a rather broad bandwidth for an entire signal.
Such a case is best exemplified in the case of the carnivorous Spear-nosed bat (Phyllostomus hastatus), who emits four harmonics (1°, 35 kHz to 30 kHz; 2°, 45 kHz to 35 kHz; 3°, 60 kHz to 45 kHz; 4°, 75 kHz to 60 kHz). This results in an overall band width of 40-45 kHz.
A second pattern involving broadband is the frequency-modulated signals. It consists of a brief FM sweep (1-3 ms), followed by a short (5-7 ms) constant-frequency component. This FM followed by short CF pattern is found in many tropical and temperate species which are often called as ‘FM bats’ (example: Big Brown bat, Eptesicus fuscus).
(b) Narrowband, constant frequency signals:
The first narrow-band CF patterns consist of multiple harmonic, short CF signals. These signals are of very short duration (0-2-2 ms) and lack any substantial FM sweep. However, they are very much like that of multiple harmonic FM signals.
The fundamental frequency is emitted at or near 20 kHz and is accompanied by four harmonics. These harmonics are all CF or only slightly FM. The acoustic orientation sounds of the Greater False vampire (Megaderma lyra) is the best example within this group.
A second narrowband CF pattern consists of a moderately short CF signal (5-7 ms) followed by a brief FM sweep (1-2 ms). There may be one to three harmonics and the FM portions of these are shallow so as to prevent overlap in bandwidths. For example, Pteronotus gymnonotus, P. davyi and P. personatus of the family Mormoopidae are of this sort.
A third CF pattern is seen similar to the second except that the CF component is exceptionally long (20 to over 100 ms) in duration. This pattern is common among the species of the old world family Rhinolophidae (Rhinolophus landeri) and some members of the Hipposideridae.
(c) Variable bandwidth:
Several insectivorous species of the genus Tadarida are quite flexible in their use of acoustic orientation patterns. These bats are perhaps the most versatile echo locators among the micro chiropterans. In uncluttered spaces, these bats hunt with narrowband short CF signals with no FM component. However, while pursuing potential prey, they drop the CF signal while adding a broadband FM component.
One harmonic accompanies this FM sweep. In cluttered or confined situations, these bats employ a multiple harmonic signal with several harmonics that overlap somewhat. The same shifting from essentially FM signals to short CF/FM signals during different phases of prey hunting has also been noted in the case of fishing bats (Noctilio leporinus).
Effectiveness of acoustic orientation of bats
(a) Target size discrimination:
Recent experimental studies have revealed that the little Brown bat (Myotis lucifugus) is capable of detecting and flying through an obstacle network of wires 0.28 mm in diameter. The phyllostomids (Glossophaga soricina, Carollia perspicillata and Artibeus jamaicensis) can detect wires ranging from 0.19 mm to 0.175 mm in diameter.
The Mediterranean Horseshoe bat (Rhinolophus euryale) and the megadermatid (Megaderma lyra) has the ability to detect wires as fine as 0.05 mm and 0.08 mm in diameter, respectively, which is about the thickness of human hair.
(b) Target range discrimination:
Target range (distance) discrimination of echo locating bats varies with the size of the target. For example, the Little Brown bat (Myotis lucifugus) can detect wires of 3 mm diameter at range of about 2.25 m. This range decreases to 1.15 m for 0.28 mm wires and 1.0 m for 0.18 mm wires.
Similarly, the Mediterranean Horseshoe bat (Rhinolophus euryale) does not detect a 3 mm wire until it is within 1.4 mm distance of the wire, and a wire of 0.05 mm thickness is perceived at a distance of 0.2 m. It has been demonstrated that bats (Eptesicus fuscus, for example) have the ability to differentiate between two identical targets (as little as 12-13 mm) placed at different distances.
(c) Target shape and texture recognition:
It has been worked out that bats can differentiate target shape and texture of objects. It has been experimentally proved that the carnivorous phyllostomid, Vampyrum spectrum, is able to detect differences in the shape of various spheres and spheroids. Demonstrations in case of the Brown bats has proved their ability of textural discrimination of plexiglass plates with as little as 0.8 mm difference in hole depth.
(d) Feeding rates:
The efficiency of bat ecolocation activity has been evaluated by using feeding rates. It has been observed that the feeding rate of individuals of Myotis lucifugus in terms of successful capture of insects in the field is 1 g hr-1.
The prey size varied between 0.2 mg and 3 mg. Thus, this species of bat detects and captures insects, in the wild, at a rate of about 500 per hour or about one insect every seven seconds. The same species, while foraging in a room full of Drosophila (about 2 mg), demonstrated a capture rate of 1,200 per hour or one every three seconds.
Working Space and Information Content of Echoes:
The important task an echo locating bat faces is to sort out objects acoustically in its immediate environment. The dimension of this ‘working space’ vary among species of bats from less than a meter to several meters, depending on flight speed and style, as well as target range discriminatory ability.
Within this working space the bat must be able to differentiate food items and avoid potential obstacles and all this has to be accomplished in a matter of microseconds.
The various echolocation patterns (multiple harmonic FM, FM/short CF, short CF/FM etc. as described above) serve as a carrier of vital information about target size, distance etc. Each of these acoustic patterns is limited by the kind of target imagery that it can convey.
(a) General Pattern of Echolocation:
A generalized sequence of acoustic events appears to be common to all echo locating bats and can be divided into three distinct phases – search, approach and terminal. Each phase of echolocation is characterised by a marked quantitative change in the rate at which the ultrasonic sounds are emitted and the duration of each signal. Fig. 5.23 illustrates the nature of these three phases in four different species of bats. Each of these bats uses a different qualitative pattern of acoustic orientation.
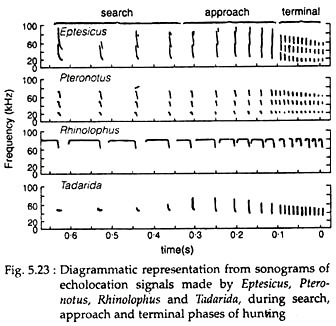
(i) Search phase:
In most bats, including those given in Fig. 5.23, the search phase is characterised by a signal emission (or pulse) rate of about 10 pulses per second. As ultrasonic sound production is a metabolically expensive process, the repetition rate of the acoustic signals is regulated or balanced against the activity of that moment.
Signals are emitted at a minimal rate, during the search phase, which is just sufficient to survey and screen the area ahead for potential prey items or obstacles. The rate of signal emission in the search phase also appears to be correlated with flight speed and the degrees of environmental chitter.
(ii) Approach and terminal phase:
Goal oriented flight begins once the target has been detected. It gives rise to the approach phase, characterised by 25-50 pulses per second. It finishes with the terminal pulse during which pulse emission may reach as high as 200 or more pulses per second.
The marked rise in signal emission rate during these two goal-oriented phases allows the bat to gather more precise information concerning the speed, direction of movement and other acoustic imagery about the intended target or obstacle. Due to the rapidity of the terminal phase, it is often termed as the “buzz”.
In many echo locating bats, although the general structure of the signal remains unchanged, the qualitative nature of each signal may be slightly modified, as in the case of Rhinolophus, where the duration of the signal is shortened.
In contrast, the Mexican Free-tailed bat (Tadarida brasiliensis) and the Long-eared vespertilionid (Idionycteris phyllotis), show a marked change in the structure of the acoustic signal during search, approach and terminal phases. During search phase they use a narrowband CF signal.
In the approach phase and while hunting in open areas, they drop the CF component and add a FM component. In complex, cluttered areas, they add multiple harmonics as well. Thus, these bats use different acoustic signals for different hunting situations.
(b) Pulse or Echo Overlap:
During the search phase of echolocation, the initial detection of a target takes place. The position of a target is first perceived when an echo, after getting reflected from the object, arrives back at the bat’s ears. The distance of the object can thus be evaluated by the time interval between a pulse emission and the return of its echo.
In some bats, the echo from a pulse arrives at the time when the next pulse emission has already taken place. The Wagner’s Moustached bat (Pteronotus personatus) emits a search pulse of about 4 ms duration. We know that sound travels through air at 340 mm ms-1, a pulse of 4 ms duration would be 1,360 mm in length.
As the pulse or echo circuit is a two-way journey, and then this species would receive an echo from an object about 680 mm ahead at just about the time when the next pulse was getting emitted. With a flight speed of 1.7 mm s-1 and a search pulse rate of 18 (4 ms) pulses per second, the next echo would return or overlap about 0.6 ms into the succeeding pulse emission. Similarly, the next echo would overlap by 0.12 ms.
However, most species of echo locating micro chiropterans appear to avoid pulse or echo overlaps. On making contact with the target, the pulse duration is shortened. The Little Brown bat (Myotis lucifugus) searches its prey at a pulse rate of about 15 per second, with each pulse being about 2.5 ms duration.
These bats can detect targets at about 2 meters distance, but do not attend to them until nearing about 720 mm. The bat, as it approaches its target, shortens the pulse by about 1 ms per 260 mm. Just before contact the pulse may be as short as 0-25 ms with the pulse rate being about 200 pulses per seconds.
(c) Narrowband Target Imagery:
Long CF pulses appear to be associated with searching and target or obstacle detection at generally long distances. Such acoustic signals may be more useful at high altitudes where sound attenuation is reduced. However, these signals are of maximal efficiency in open, uncluttered situations.
As shown in Fig. 5.24A, short CF signals are relatively limited in the information content they can carry. They are good only for the initial detection of a target, but do not convey any other information than the general location of the target.
They are of value mainly during close pursuit of prey, flight in congested conditions and perhaps also in landing. It is, thus, not surprising that there are no bats that use such type of signals exclusively.
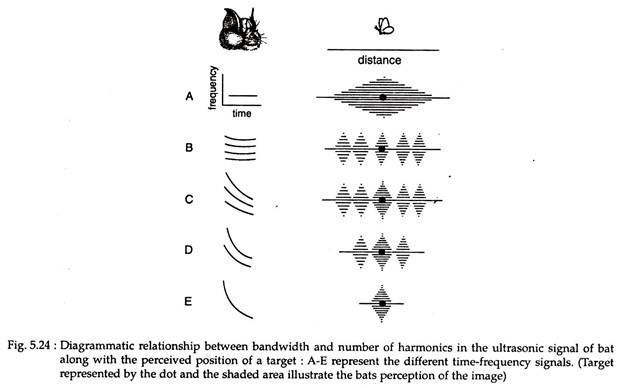
Adding several harmonies to the short CF signal (Fig. 5.24B) sometimes improves somewhat the perception of the target’s ‘true’ position. Such crude type of acoustic orientation appears to be used by Megmaderma lyra, rhinopomatids and nycterids.
(d) Broadband Target Imagery:
Bats that move about and forage in complex, cluttered environments use multiple harmonic signals with broadband FM sweeps (Fig. 5.24C). In many bats, the overall bandwidth is large (Fig. 5.24D), while others may use but a single broadband, curvilinear FM sweep (Fig. 5.24E).
However, all FM signals provide better multidimensional acoustic images than do CF signals. Better localisation of a target’s ‘true’ position increases as bandwidth is broadened. This has been amply illustrated in Fig. 5.24C-E.
Use of multiple harmonics and its correlation with the hunting situation can be best exemplified in the case of the genus Myotis. Myotis volcans hunt over the open fields and clearings and uses one prominent harmonic, while Myotis evotis uses narrow fundamental frequency and two harmonics to hunt in tangled vegetation.
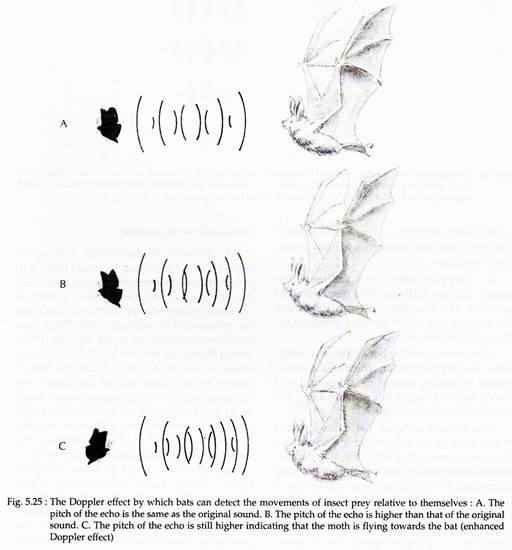
(e) Doppler Effect:
Different bats use different methods of echolocation. It can determine the distance to prey by the time required for the signal to bounce back. The echoes coming back from any insect show the Doppler Effect, which is, if a sound source is moving toward us, the sound will have a higher pitch; if it is moving away, the sound will be of lower pitch.
The horseshoe-nosed bat emits a signal at 83-4 kHz. If the echoes coming back from the insect (prey) are at 83-4 kHz, they tell the horseshoe-nosed bat that the insect is flying away from it at the same speed that the bat is travelling (Fig. 5.25).
If the sound comes back lower than 83-4 kHz, the insect is moving away faster than the bat; and if the sound comes back at a higher speed then it means that the bat is closing on the insect. The bat then lowers the frequency of its signal.