
In this article we will discuss about:- 1. Introduction to Metamorphosis in Amphibians 2. Causation of Metamorphosis in Amphibians 3. Tissue Reactivity 4. Induction.
Introduction to Metamorphosis in Amphibians:
In amphibians, metamorphosis is associated in typical cases with a transition from an aquatic to a terrestrial mode of life. Super-imposed on this change in environment, there is in the anurans (frogs and toads) a change in feeding. The tadpoles of most frogs and toads feed on vegetable matter—particles of plants, living and decaying—which they scrape off submerged objects with the aid of the horny teeth surrounding their mouths.
Some are detritus feeders, passing through their guts the mud and detritus collected from the bottom, and others, such as the tadpole of Xenopus, are plankton feeders. Adult frogs are carnivorous, living on insects, worms, and the like, but sometimes also on larger prey, such as smaller frogs and even little birds and rodents, which they catch, overpower, and swallow. In the case of urodeles, there is no substantial change in diet, the larvae being as carnivorous as the adults, though naturally feeding on smaller animals (mainly crustaceans and worms).
The changes in the organization of the animals during metamorphosis are in part progressive and in part regressive and may be grouped into three categories:
ADVERTISEMENTS:
1. The organs or structures necessary during larval life but redundant in the adults are reduced and may disappear completely.
2. Some organs develop and become functional only during and after metamorphosis.
3. A third group of structures, while present and functional both before and after metamorphosis, become changed in order to meet the requirement of the adult mode of life.
In anurans, the differences between the modes of life of the larva and the adult are much more profound, and accordingly the changes at metamorphosis are more extensive than in the urodeles. We will consider them first.
ADVERTISEMENTS:
The regressive processes occurring during the metamorphosis in frogs are the following:
The long tail of the tadpole with the fin folds is resorbed and disappears without a trace. The gills are resorbed, the gill clefts are closed, and the peribranchial cavities disappear. The horny teeth of the perioral disc are shed as well as the horny lining of the jaws, and the shape of the mouth changes. The cloacal tube becomes shortened and reduced. Some blood vessels are reduced, including parts of the aortic arches.
The constructive processes involve, first, the progressive development of the limbs, which increase in size and differentiation. The forelimbs, which in the frogs develop under cover of the opercula membrane, break through to the exterior.
The middle ear develops in connection with the first pharyngeal pouch (the pouch situated between the mandibular and the hyoid arches). The tympanic membrane develops, supported by the circular tympanic cartilage. The eyes protrude on the dorsal surface of the head and develop eyelids. The tongue is developed from the floor of the mouth.
ADVERTISEMENTS:
The organs which function both in the larva and the adult, but change their differentiation during metamorphosis, are primarily the skin and the intestine. The skin of the tadpole is covered with a double-layered epidermis. During metamorphosis, the number of layers of cells in the epidermis increases, and the surface layers become cornified. Multicellular mucous and serous glands develop as pockets sinking from the surface into the subcutaneous connective tissue layer.
The lateral line sense organs, present in the skin of tadpoles, disappear during metamorphosis in most frogs. The pigmentation of the skin is changed; new patterns and colors appear. The intestine, which is very long in tadpoles, as in most herbivorous animals, becomes greatly foreshortened, and most of the coils which it forms in the tadpole become straightened out. The metamorphosis is very rapid and takes only a few days.
In urodeles, the changes at metamorphosis are far less striking. The tail is retained; only the fin fold disappears. The branchial apparatus is reduced, the external gills become resorbed, and the gill clefts close. The visceral skeleton becomes greatly reduced. The head changes its shape, becoming more oval. The progressive changes are less conspicuous than in the metamorphosing frog tadpoles. They are restricted mainly to changes in the structure of the skin and the eyes.
The eyes bulge more on the dorsal surface of the head and develop lids. The skin becomes cornified, and multicellular skin glands become differentiated. The pigmentation of the skin changes. The legs, contrary to those of tadpoles, undergo hardly any change at all, and the same may be said of the alimentary canal. The metamorphosis is, ort the whole, more gradual and may take up to several weeks.
It is worth noting that the lungs do not undergo drastic changes during metamorphosis both in anurans and urodeles. They develop very gradually and become fully functional in the larval state. Long before metamorphosis, the larvae of frogs as well as of salamanders start coming up to the surface and gulping air into their lungs and thus supplementing their aquatic respiration. This may be of considerable importance where the larvae develop in stagnant and polluted waters, as is often the case.
Hand in hand with morphological changes during metamorphosis go physiological changes, of which we shall point out the following. In frog tadpoles, the endocrine function of the pancreas starts at metamorphosis and this is connected with the increased role of the liver in the turnover of carbohydrates (glycogen).
A profound change takes place in the excretory mechanism. In the tadpole, the end product of nitrogen metabolism is ammonia, which is easily disposed of (by diffusion) in an aquatic medium but which in a terrestrial animal might accumulate and become dangerous because of its high toxicity.
Metamorphosed frogs, however, excrete most of their nitrogen in the form of urea and only small amounts as ammonia. The changeover occurs in the late stages of metamorphosis and is, of course, due to a changed function of the liver, which performs the synthesis of urea.
The reduction of the gills and the tail in tadpoles is effected by autolysis of the component tissues of these organs, with active participation of ameboid macrophages which phagocytose the debris of the disintegrating cells. The same mechanism, though of a limited scope, is in action when the external gills and fin folds are reduced in urodele amphibians.
ADVERTISEMENTS:
Since destructive processes play such a considerable role in metamorphosis and since, in addition, food intake may be interrupted during the crucial part of the transformation, especially in tadpoles, the mass of the body at the end of metamorphosis is smaller than at the beginning (“de-growth,”).
The reduction of the body mass is due not only to loss of some parts (gills, tail), but the remaining parts, excepting the actively growing organs, also appear to shrink during metamorphosis – the head and trunk of metamorphosed amphibians are smaller than in larvae just before the commencement of metamorphosis.
Causation of Metamorphosis in Amphibians:
The concurrent changes in so many parts of the animal’s body during metamorphosis suggest the existence of some common cause for all the transformations. It has been found that this common cause is a hormone released in large quantities from the thyroid gland of the animals entering the stage of metamorphosis.
The first indication of this was obtained when Gudematsch fed frog tadpoles on dried and powdered sheep thyroid gland and observed that they metamorphosed precociously (1912). Feeding tadpoles with preparations of other glands did not have the same effect. This experiment made it seem very probable that tadpoles are capable of reacting to the thyroid hormone by metamorphosing.
That the thyroid hormone is actually the cause of metamorphosis in normal development was further proved by the following two experiments. The rudiment of the thyroid gland was removed in frog embryos in the tail-bud stage (through an incision on the ventral side). The operated tadpoles were fully viable and showed normal growth but failed to metamorphose, although they were kept alive almost a year after the control animals had become little froglets.
The thyroid-less tadpoles continued to grow and attained a much greater size than normal, having a total length of up to 123 mm. instead of about 66 mm., as at the beginning of metamorphosis in normal tadpoles. It was thus proved that metamorphosis cannot set in without a stimulus emanating from the thyroid gland. The final experiment consisted in supplying the thyroid-less tadpoles with thyroid hormone from without, either by feeding them on dried thyroid gland or by immersing them in water containing soluble extracts from thyroid glands.
The tadpoles treated in this way immediately proceeded to metamorphose, thus showing that their own thyroid glands were not necessary as long as they were supplied in some way or other with thyroid hormone.
Similar experiments were carried out on urodeles. A very suitable animal for the experiments is the axolotl Ambystoma mexicanum, which under ordinary conditions does not metamorphose at all but may be induced to metamorphose by thyroid treatment.
The active principle of the thyroid gland may be introduced into the animal’s body in several different ways. Normally it emanates from the animal’s own thyroid gland. The same effect may be produced by implanting bits of live thyroid gland, feeding the animals on thyroid gland, injecting them with preparations of thyroid gland, or keeping them in water containing soluble extracts of thyroid glands. The last fact clearly shows that the active principle of the thyroid gland is a chemical substance—a hormone.
A saline extract of fresh thyroid tissue contains protein, thyroglobulin, which retains the activity of the thyroid gland. An important characteristic of thyroglobulin is that it contains iodine, which, as we will see, is of great importance for the working of the thyroid hormone.
Thyroglobulin has a molecular weight of about 675,000. Its molecules are thus very large, and it is unlikely that thyroglobulin can penetrate as such through cellular membranes, which would be necessary if it were to leave the thyroid gland and reach the cells eventually reacting to the thyroid treatment.
To become active, the iodine-containing compounds are released from their connection to the thyroglobulin. Several compounds of smaller molecular weight are liberated in this way. In chemical structure, all these compounds are combinations of molecules of the amino acid tyrosine with one or more atoms of iodine. The most important of them are tri-iodothyronine and thyroxine. In both of these compounds two residues of tyrosine are joined together, and to these are attached three (in tri-iodothyronine) or four (in thyroxine) atoms of iodine.
The structural formulas of these substances follow:
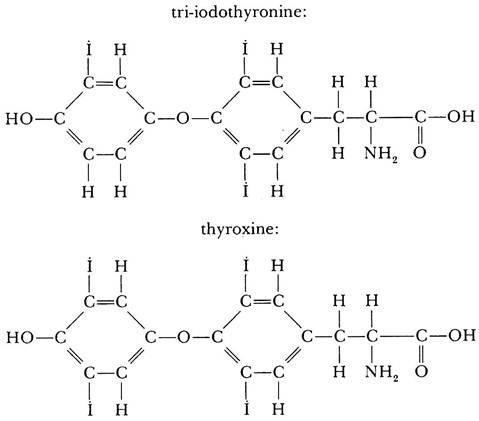
Of the two compounds, thyroxine is produced in very much greater quantities and is thus the main active substance released by the thyroid gland, although tri-iodothyronine is more highly active on tissues.
It has been tested whether iodine alone can cause metamorphosis in amphibians, and positive results have been obtained either by keeping frog tadpoles or newt larvae in water containing the element iodine (in solutions as weak as 0.0000003833 per cent!), by injecting animals with iodine, or by implanting iodine crystals into the body cavity. The element iodine caused metamorphosis even in thyroidectomized axolotls; thus, it must have been acting directly and not through the increased production of hormone by the animal’s own thyroid gland.
On the other hand, it appears that the degree of activity of the iodine atoms may be greatly influenced by the type of amino acid with which the iodine is bound. This can be clearly shown by comparing the activity of the two amino acids di-iodotyrosine and thyroxine.
By placing tadpoles in solutions of each amino acid separately, it was found that the same amount of iodine was 300 times more active when forming a part of the thyroxine molecule than when incorporated into the di-iodotyrosine molecule. The activity of tri-iodothyronine is three to five times as high as that of thyroxine.
The thyroid gland is not the only gland which is involved in the causation of metamorphosis in amphibians. It has been discovered that the hypophysis plays an important part as well. If, in frog tadpoles, the hypophysis is destroyed or if the rudiment of the hypophysis in a late frog embryo is excised, the tadpoles do not metamorphose, just as if their thyroid glands had been removed.
The removal of the hypophysis rudiment may later be compensated for by implanting pieces of hypophysis from metamorphosed or adult frogs, but only if the thyroid gland of the animal is intact. If the thyroid gland has been removed, no amount of implanted hypophyseal tissue can induce metamorphosis. From this it may be concluded that the hypophysis does not act upon the tissues directly but acts only by way of stimulating the thyroid gland.
In fact, the thyroid gland of hypophysectomized animals remains underdeveloped and does not accumulate the thyroid hormone (in the form of “colloid”) in its follicles. Implantations of active hypophysis can also be used to stimulate metamorphosis in the axolotl. The agent necessary for activating the thyroid gland is produced in the anterior (epidermal) lobe of the hypophysis and has been isolated in the form of a thyrotropic hormone.
In the case of larval amphibians, the hypophysis does not produce the thyrotropic hormone until the time when metamorphosis normally occurs. This has been proved by taking hypophyses from tadpoles of various ages and transplanting them into tadpoles whose own hypophyses had been removed previously.
Whereas hypophyses taken from tadpoles in stages of metamorphosis or from metamorphosed frogs compensated for the removal of the animals own hypophysis and restored the tadpole’s ability to metamorphose, hypophyses taken from younger stages were not effective.
The hypophysis of tadpoles also secretes another hormone which acts as an antagonist of thyroxine during the larval life of the tadpole. The hormone stimulates growth and retards metamorphosis. The chemical nature of the hormone is very similar to or possibly even identical with prolactin. In mammals, prolactin causes growth of milk glands and milk secretion. It is quite remarkable that the same or a very similar chemical substance is present in such different animals as mammals and amphibians but is used to control entirely different processes.
The initial signal for metamorphosis is given by the anterior lobe of the hypophysis when it reaches a certain degree of differentiation and becomes capable of producing the thyrotropic hormone. The thyrotropic hormone activates the thyroid gland, which synthesizes and released the thyroid hormone.
The thyroid hormone (of which thyroxine is the most important component) overcomes the action of the prolactin-like hormone and affects the tissues directly, causing the degeneration and necrosis of some cells and stimulating the growth and differentiation of others.
Tissue Reactivity in Amphibian Metamorphosis:
One of the most intriguing aspects of amphibian metamorphosis is the diverse nature of reactions in different tissues to one and the same condition—the presence of the thyroid hormone.
Whereas some of the tissues (the tail, the gills) become necrotic owing to the action of the hormone, others (the limbs) react to the hormone by increased growth and progressive differentiation. It can easily be shown that the character of the reaction is not due to the position of the parts in question, nor to an uneven distribution of the active principle, but solely to the nature of the reacting part.
Parts of a tadpole’s tail transplanted to the trunk undergo metamorphosis together with the host’s tail and become absorbed. On the other hand, an eye transplanted to the tail of a tadpole before metamorphosis remains healthy, while all the surrounding and underlying tissues undergo necrosis. As the tail shrinks, the eye is brought nearer to the trunk and eventually becomes fused to the body in the sacral region after the entire tail has disappeared.
The same experiments, incidentally, prove that the stimulus of the thyroid gland is carried through the blood vascular system, as it is only by this route that it can reach any part independently of its actual position. The secretion of the thyroid gland is thus a hormone in the narrow sense of the term.
What happens to a tissue under the influence of the thyroid hormone is determined by the reactive properties of the tissue itself, or by what we have earlier called its competence. The competence of tissues to react to the thyroid hormone is not directly dependent on their histological differentiation. In tadpoles, while the myotomes of the tail become resorbed during differentiation, the myotomes of the trunk are not so affected.
Furthermore it has been noticed that different parts of the body reacting to the thyroid hormone (whether by degeneration or progressive development) are not equally responsive to the dosage of the hormone. Very weak dosages applied to frog tadpoles cause an acceleration of the growth and differentiation of hind-limbs and a shortening of the intestine.
Further processes may not be set into motion at all or follow only after a lengthy treatment. A higher dose of the thyroid hormone causes the breakthrough of the forelegs. An even greater dose of the hormone is necessary to cause the resorption of the tail.
There is evidently some kind of threshold value for each part which has to be attained before the reaction sets in. Different parts of the tail have different threshold values, the tip of the tail reacting more readily than the proximal parts. In general, the degree of sensitivity to the thyroid hormone is reflected in the order in which the metamorphic changes proceed in normal development – the parts which have a low threshold (legs, reacting by growth) respond earlier than the parts having a high threshold value (tail, reacting by reduction).
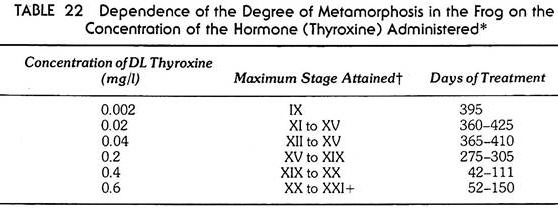
The importance of the dose of hormone is evident from Table 22, which shows how far metamorphosis can proceed depending on the amount of hormone being administered. When heavy doses of thyroid hormone are supplied to young tadpoles, all processes start at once and the normal sequence of events become upset, the destructive processes being capable of proceeding faster than the constructive processes. The forelimbs break through before becoming differentiated; the tail becomes reduced before the legs are sufficiently developed to take over locomotion. The result is, of course, the death of the animal.
In urodeles, the bulging of the eyes seems to be the reaction which may be elicited by the weakest doses of the thyroid hormone. Next follow, more or less concurrently, the reduction of the fin fold and the shortening and disappearance of the external gills.
The closure of the gill clefts and the transformation of the skin are the results of maximal stimulation and accompany complete metamorphosis in the course of normal development. The earlier stages of metamorphosis, including the shortening of the external gills, are partly reversible- if the thyroid treatment is stopped, the gills may elongate again to a certain extent.
The influence of thyroxine on metamorphosis in amphibians is exercised by way of a modification of the activity of the genome in the reacting tissues. This has been proved not so much in respect to the morphological transformations during metamorphosis, but rather in respect to a physiological change—a change in the biochemistry of excretion, which as we have seen accompanies metamorphosis in frogs. At metamorphosis tadpoles change from excreting nitrogen waste in the form of ammonia, to excreting it as urea.
The urea is produced in the liver of the animals by a system of enzymes linked together in the so-called “ornithine urea cycle.” The rate- limiting enzyme of this cycle is the enzyme carbamyl phosphate synthetase (CPS). It has been found that the production of the enzyme can be significantly stimulated by thyroxine at a concentration of 2.6 x 10–8 mol. This stimulation is sensitive to actinomycin, which inhibits the production of the enzyme if added to an in vitro test system consisting of cubes of tadpole liver tissue, with thyroxine in the culture medium.
As actinomycin is a transcription inhibitor, it follows that thyroxine stimulates production of CPS (and thus activates the ornithine urea cycle, and the manufacture of urea) by stimulating the transcription of mRNA for carbamyl phosphate synthetase.
Lastly, it was shown that the first reaction to thyroxine is an increase of nuclear RNA in the tissues of tadpoles. This reaction is observed within 50 hours of the application of thyroxine. About 25-30 hours later there is an increase in cytoplasmic RNA, and the increase in carbamyl phosphate synthetase is observed only about three days later.
Induction during Amphibian Metamorphosis:
Although, as a general rule, the processes of metamorphosis are a direct reaction to the thyroid hormone which reaches each tissue, there are some notable exceptions to this rule. The skin covering the tail of a tadpole, although subject to necrosis in normal metamorphosis, remains healthy if transplanted to the body, provided that it is transplanted without the underlying muscles. Skin transplanted with underlying tail muscle will necrotize in any position on the body.
It is thus evident that the direct action of the thyroid hormone is on the muscle tissue, and the overlying skin becomes involved in the process of resorption secondarily. A more complicated case is presented by the development of the tympanic membrane in frogs. The middle ear, with its cavity connected by the eustachian tube to the pharynx, is one of the structures that develop progressively during metamorphosis. The tympanic membrane first becomes differentiated toward the end of metamorphosis.
It is supported in frogs by a cartilaginous ring, the tympanic cartilage, which develops as an outgrowth from the posterior edge of the quadrate cartilage. The skin which later participates in the formation of the tympanic membrane is originally no different from the skin covering the rest of the body.
During metamorphosis, the connective tissue layer of the skin in the area of the tympanic membrane becomes reorganized, the original layer of fibers, the stratum compactum, is broken up with the participation of phagocytes, and a completely new, somewhat thinner fibrous layer is developed in its place. In the fully differentiated tympanic membrane, the skin is less than half the thickness of ordinary skin but much more compact, and it also differs in its pigmentation.
It has been found that the differentiation of the tympanic membrane is not due to a direct action of the thyroid hormone but that it is induced by the tympanic cartilage. If the tympanic cartilage is removed before metamorphosis, the tympanic membrane does not develop.
If the area in the otic region is covered by skin from the flank or from the back, the skin will develop a tympanic membrane. Lastly, if the tympanic cartilage is inserted under the skin on the flank or the back of a tadpole approaching metamorphosis, the local skin becomes differentiated as tympanic membrane.
We can draw up a complicated chain of interactions that must take place before a tympanic membrane becomes differentiated. The first step is the formation of the rudiment of the hypophysis. The latter is developed in conjunction with the stomodeal invagination and presumably is induced by the oral endoderm.
The hypophysis in due time secretes the thyrotropic hormone and activates the thyroid gland. The thyroid gland releases the thyroid hormone, which causes the posterior edge of the quadrate to become differentiated as the tympanic cartilage. Lastly, the tympanic cartilage induces the skin to differentiate as tympanic membrane.