
In this article we will discuss about:- 1. Meaning of Speciation 2. Rate of the Speciation Process 3. Mechanism 4. Classification 5. Modes 6. Rate.
Definition of Speciation:
Species has been considered to be the product of the opportunistic process of evolution. Taxonomist encounters difficulties in trying to delimit species mainly due to two aspects of evolution: variability within populations and the existence of incipient species.
The formation of new species is referred to as speciation. There are different attitudes towards speciation. Some authors even deny the existence of speciation as a distinct natural phenomenon. For many biologists, every speciation event has its individuality, being the outcome of a unique mixture of different ingredients, such as geographical context, and the number and type of genetic structures involved.
For others, there is a very limited range of fundamentally different modes of speciation. While, a few researchers believe that speciation involving sexually reproducing organisms are fundamentally the same. These different attitudes reflect the uneven attention paid to recognise several different and partly independent aspects of speciation.
ADVERTISEMENTS:
Is speciation an adaptive process? This is perhaps one of the most basic of the many questions regarding the causes of speciation. The most prevalent view is, perhaps, expressed by Herman Muller (1940), Ernst Mayr (1963) and many later authors.
According to them “reproductive isolation evolves as a by-product of genetic changes that occur for other reasons, so that speciation is an incidental, non-adaptive consequence of divergence of populations”.
Rate of the Speciation Process:
When considering the rate of the process, two modes of speciation can be distinguished:
(i) Conventional, gradual speciation occurring in the course of a long series of generations, and
ADVERTISEMENTS:
(ii) Instantaneous speciation.
In population genetic terms, Templeton (1982) similarly distinguished between the slow speciation by divergence and the quick speciation by transilience (Table 3.3B).
Mechanism of Speciation:
To understand the mechanism of speciation, one must have a clear knowledge of biological species concept. A species is considered to be a group of actually or potentially interbreed natural population that is reproductively isolated from other such groups.
Thus, the individuals of a species interbreeds with one another, but no interbreeding occurs between two different species. As a result of interbreeding, gene flow occurs either within the population or between populations (Fig. 3.2).

Gene flow occurs between members of the same species; but no gene flow occurs between two different species. Each species maintains its integrity as long as its gene pool does not mix with that of other species. The mixing of gene pool will not be possible without breeding of interspecies. Such interbreeding is not possible when there are genetic differences between the two groups. Species, thus, evolve owing to genetic divergence and isolating mechanisms.
If a single randomly mating population gives rise to two reproductively isolate populations, then it is generally not sufficient for a single mutation to confer reproductive isolation on its bearers. Such type of mutation (say, X2) may arise in a single or a few individuals and in heterozygous condition (X1X2).
Its reproductive fitness will be lowered, irrespective of the fact whether it takes place in post-zygotic isolation (example, sterility) or pre-zygotic isolation (example, failure to mate with the ‘normal’ type). Therefore, it will generally be eliminated by natural selection.
In the case of polygenic trait as a cause of reproductive isolation, the problem is that recombination generates intermediates. Thus, the problem of speciation is how two different populations can be formed without intermediates.
There are various controversies concerning speciation. These are due to the fact that the opponents did not see clearly that speciation has two aspects — genetic and populational. These two factors are not naturally exclusive alternatives; rather, both are always involved simultaneously.
Classification of Speciation:
The modes of speciation have been classified by two authors separately (Table 3.3). The one classified by Mayr (1963) is generally followed.
The evolution of reproductive barrier based on several or many allele substitutions has been referred by Mayr as gradual speciation. Such gradual speciation may occur under three geographic settings— allopatric speciation, parapatric speciation, and sympatric speciation.
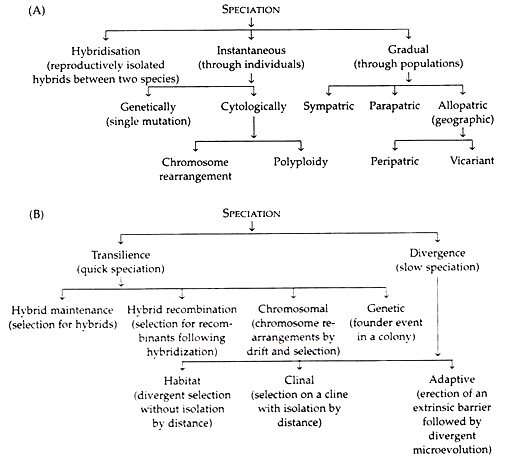
Other than the above-mentioned ones, some instances are also known where species have arisen from hybrids. Here speciation occurs chiefly by polyploidy or even by chromosomal rearrangement.
Modes of Speciation:
ADVERTISEMENTS:
A. Allopatric Speciation:
When two related populations are geographically separated by an extrinsic barrier, physical barrier (such as topography, water or land) or un-favourable habitat, it results in the evolution of gradual genetic reproductive barriers between them. This results in the formation of separate species and the process is called allopatric speciation.
At the beginning the gene pool of the two related populations (allopatric population) is similar. Due to the formation of the geographic barrier, there is no possibility for the mixing up of genes between the populations. The environmental factors act upon the two populations differently. The two populations are under the operation of natural selection and genetic drift — which leads to genetic divergence.
When genetic divergence has developed, the populations, which once made effective interbreeding, fail to interbreed when they are brought together by migration or by the disappearance of the barrier. Thus, reproductive isolation has come into existence and the two populations are now considered to be separate species.
In species that disperse little or are strongly tied to a particular habitat, extrinsic barrier may isolate populations on a “micro- geographic” scale, such as segregated habitats within a lake. Such extrinsic barriers need not reduce gene exchange to zero, but may reduce to a very low level. All evolutionary biologists are of the opinion that allopatric speciation does occur in nature and is the prevalent mode of speciation in animals.
Kinds of Allopatric Speciation:
Two major models of allopatric speciation have been postulated, differing in population structure and genetic dynamics:
(a) Vicariant Speciation:
Vicariance occurs when two rather widespread populations are divided either by the emergence of an extrinsic barrier, or the extinction of intervening populations, or migration into a separate region (Fig. 3.3A). One such example is the emergence of the Isthmus of Panama during the Pliocene.
It divided many marine organisms into Pacific and Caribbean populations, some of which have diverged into distinct species. Vicariant speciation is supposed to occur by the operation of natural selection and perhaps by genetic drift, in both the populations separately and, perhaps, to a greater extent in one than the other.

(b) Peripatric Speciation:
Peripatric speciation is another major mode of allopatric speciation. It occurs when a colony that is derived from a much widespread parent population diverges and acquires reproductive isolation (Fig. 3.3B). This model, sometimes called founder-effect speciation, is a very controversial one, as a great deal of controversy surrounds the genetic changes postulated for such populations. Many population genetists believe that the genetic changes underlying peripatric speciation are the same as those in vicariant speciation.
According to Mayr (1954, 1963), Templeton (1980) and others, this process is thought of as speciation due to a shift, initiated by genetic drift and followed by natural selection between adaptive peaks. It has been seen in many birds and animals that isolated populations with restricted distributions often are highly divergent, even though their ecological environment appears similar to that of the parent.
Mayr explained such divergence, proposing that:
(1) Genetic change could be very rapid in localised populations founded by a few individuals that are cut off from gene exchange with the main body of the species. Differences in ecological selection plays a vital role because the environment of a small area (where the founder species are) is often more homogeneous than that of a large area. Therefore, the conflicting pressures that act on a widespread population is less numerous here.
(2) Allele frequencies at some loci differ from those in the parent population because of accidents of sampling (that is genetic drift), as a few colonists (the founders) will carry only some of the alleles from the source population, and at altered frequencies (such initial alteration of allel frequencies led Mayr to coin the term founder effect).
As epistatic interactions among genes affect fitness, this initial change in allele frequencies at some loci alters the selective value of genotypes at other interacting loci. Thus, the “genetic environment” of the other loci is altered.
(3) As the colony is cut off from the gene flow among populations the genetic environment of the colony, that imposes selection on alleles (including new mutations) is less variable, more homozygous and may favour alleles that confer high fitness in specific combinations with certain alleles at other loci.
According to Mayr (1963) the epistatic interactions are so strong that each genetic change might trigger others, so that massive genetic change might take place, yielding reproductive isolation as a by-product. Such substantial evolution might occur so rapidly and/or so localised a geographic scale, that it will seldom be documented in the fossil record. This hypothesis may thus help to explain the rarity of fossilized transitional forms among species.
An example of founder-effect speciation is the robin, Petrocia multi-colour. The bright plumage of the male is quite contrasting with the duller plumage of the female throughout eastern Australia. But among the various Melanesian islands present nearby, the male robins have “female” colouration on some, while the females have “male” colouration on others.
Another example of peripatric speciation is the kingfisher, Tanysiptera galeata. It shows only slight variation in plumage throughout climatically very different regions of New Guinea. However, it has given rise to several markedly different forms on the various small islands present nearby.
B. Parapatric Speciation:
Parapatric speciation is the evolution of reproductive isolation between populations that are continuously distributed in space, so that there is substantial movement of individuals and, hence, gene flow between them.
Here the populations of a widespread species diverge by adaptation to different environments. Divergent selection—even at a narrow environmental discontinuity—may oppose gene flow and result in reproductive isolation.
The importance of parapatric speciation is a matter of dispute mainly because the empirical evidence is usually ambiguous. It is a matter of question as to how reproductive isolation develops in the face of gene exchange. A probable answer is that divergent selection for the different gene combinations may be stronger than the rate of gene flow.
If gene flow is low, as probability exists over a very wide area, then relatively weak selection suffices to establish clines at the relevant loci. If gene flow is appreciable, as happens between neighbouring populations, then, due to strong divergent selection, a steep cline is formed. To say that divergent selection is strong then it has to be presumed that there exists a barrier to gene exchange.
This barrier is caused due to reproductive failure of individuals with the “wrong” genotype or phenotype that migrate across the border. Therefore, in principle, strong divergent selection at certain loci, causing a steep cline, acts like a physical barrier, resulting in genetic diversity at other more weakly selected loci. Subsequently, clines at various loci may tend to develop at the same location, resulting in a hybrid zone.

The best possible example of parapatric speciation is Authoxanthum adoratum, one of several grasses that evolved tolerance to heavy metals in the vicinity of mines. Several populations, under very strong selection for heavy metal tolerance, have diverged from neighbouring non-tolerant populations, not only in tolerance but also in flowering time.
Moreover, they self-pollinate more frequently, as they have become more self-compatible. Both these characteristics provide sufficient reproductive isolation from adjacent non-tolerant genotypes.
C. Sympatric Speciation:
Sympatric speciation is the evolution of reproductive isolation within a randomly mating population (Fig. 3.5). As the two populations inhabit the same place and in the same habitat, they have the same needs, with regard to food, space and other requirements of life. This competition brings about the operation of Gause’s law or competitive exclusion principle, which states that two populations cannot continue to occupy the same habitat indefinitely.

Sympatric speciation is one of the most controversial subjects in evolutionary biology. Sympatric speciation would arise if a biological barrier to gene exchange arose within the confines of panmictic (randomly mating) population or if speciation occurs despite high initial gene flow.
Many authors put forward that spatial segregation was necessary for speciation. Mayr (1982) demonstrated that many supposed cases are unconvincing and that the sympatric speciation hypothesis must overcome severe theoretical difficulties.
However, the one mode of sympatric speciation that is above any controversy is speciation by polyploidy in plants. Here, a single instantaneous change reproductively isolates a new polyploid from its ancestor. Any model of sympatric speciation must overcome the difficulty of how to reduce the frequency of the intermediate genotypes that would act as a channel for gene exchange.
Possible examples of sympatric speciation:
Some authors are of the opinion that sympatric speciation are quite frequent in certain groups of organisms.
A few examples are:
1. Treehoppers, Enchenopa binotata, consist of six sympatric sibling species, each restricted to a different genus of host plant. As their eggs hatch at different times, they reach reproductive maturity and mate at somewhat different times during the summer.
Wood and Keese (1990) suggested that gene flow among the species was initially reduced by differences in the timing of their life history caused directly by the host plants and that this reduction allowed subsequent genetic divergence in host preference and mate preference.
2. The apple maggot fly, Rhagoletis pomonella, shows genetic divergence due to habitat isolation and is undergoing the process of sympatric speciation. Subsequent studies by Feder (1993) have shown this to be one of the few convincing cases of incipient sympatric speciation.
D. Quantum Speciation:
Grant (1971) defined quantum speciation as the budding off of a new and very different daughter species from a semi-isolated peripheral population of the ancestral species. Quantum speciation (Fig. 3.6) is a more rapid and abrupt mode of species formation.

This type of speciation is based on the observation of Drosophila inhabiting Hawaii islands. These islands are not older than 7 million years. Nevertheless, they contain about 700 species of Drosophila, and each of these species have no sub-species.
Hawaii islands are said to be characterised by volcanic eruptions. These volcanic eruptions kill the pre-existing flora and fauna. Occasionally gravid female flies are carried to this area affected by volcanoes. In such a habitat, the fly population increases rapidly. Since this area is free from predators, natural selection is relaxed temporarily and this leads to the development of new genetic variation.
As the area becomes thickly populated, this expansion may be followed by a population crash, which will eliminate almost all the individuals. A few flies that recover from the crash are acted upon by natural selection and genetic drift. This leads to the formation of genetic divergence and the failure of interbreeding with the original population by the development of reproductive isolation.
E. Polyploidy and Hybrid Speciation:
(a) Polyploidy:
These are special considerations that play a greater role in diversification and speciation in plants than in animals. Speciation by polyploidy has two distinctions:
1. It is the only known mode of instantaneous speciation by a single genetic event.
2. It is the only mode of sympatric speciation.
Among animals, polyploidy is rare. Sexually reproducing polyploids are known in a few groups of fishes (particularly in trout and sucker fishes) and frogs. Polyploid animals are often parthenogenetic, such as weevils, grasshoppers, salamanders and lizards. Polyploidy, however, is very common in plants.
Polyploidy generally occurs due to failure of the reduction division in meiosis yielding 2n gametes. Union of such a gamete with haploid (normal) or diploid gametes result in triploid (3n) and tetraploid (4n) zygotes, respectively.
(b) Hybridization:
Hybridization may sometimes give rise to distinct species. In plants, some genotypes of diploid hybrids are fertile and are reproductively isolated from the parent species, and so give rise to new species. It may increase in frequency, forming a distinct population. Verne Grant (1971) named such speciation recombinational speciation.
Several researchers have produced “hybrid species” of plants experimentally through independent translocations, such that parts of chromosomes A and B of species 1 are reciprocally trans-located in species 2. Similar reciprocal translocations are done for other chromosomes.
Although, in most cases, crosses of such types result in largely sterile progeny, nevertheless, an F2 that is homozygous for one of these combinations will form fertile hybrids. One such example is the experiment of Grant (1966) who crossed Gilia malior with G. mudocensis — resulting in 99% sterile F1. However, he was able to obtain several F2 plants and reared inbred lines by self-fertilization.
Although such hybridization may be a common mode of speciation in plants, it is rare in animals. One such possible example, described by Bullini (1994), is the minnow, Gila seminuda, of Western North America.
Rates of Speciation:
The rate of speciation is related to two concepts:
(a) The rapidity with which populations of a species evolve reproductive isolation.
(b) The numbers of species that arise per unit of time. The rate of increase in the number of species depends on both the rate of origin and the rate of extinction of species.
The time required for speciation or in other words for the evolution of reproductive isolation is highly variable. However, geological evidence, combined with estimates of divergence time based on molecular data, suggests that generally the time required for full reproductive isolation is about 3 million years.
Factors affecting speciation rate:
The factors affecting speciation rate are generally:
1. Abundant topographic barriers that provide opportunities for allopatric divergence;
2. Low dispersal rates that reduce gene flow among populations;
3. Strong sexual selection, which may cause divergence in mate recognition systems;
4. Probably ecological specialisation, which may limit species to isolated patches of habitat and afford the opportunity for divergence;
5. Perhaps bottlenecks in population size, which may facilitate genetic peak shifts or substitution of chromosome rearrangements, and
6. Genetic properties (example, degree of epistasis) that may affect the rate of evolution of reproductive incompatibility (these are, however, poorly understood).