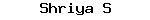
Here is a List of receptors and sense organs found in animals.
Invertebrate Receptors:
1. Mechanoreceptors:
Four types of mechanoreceptors are present in invertebrates:
i. Tactile receptors:
The tactile (touch) receptor is a phasic receptor. In cnidaria the tactile receptors cnidocils are present (Fig. 14.1) in the cnidocytes (cnidoblasts). The cnidocil being touched, the cnidocyte contracts violently discharging the nematocyst. This effects both defence and capture of prey.
ADVERTISEMENTS:
Sensory cells are scattered on the body surface of turbellaria, earthworm; on the tentacles and palpi of Neanthes, on the antennae and palpi in arthropods and on the tentacles, labial palps, head and visceral stalk in molluscs.

ii. Proprioreceptors:
The proprioreceptors are tonic receptors, and register stress changes in the exoskeleton. In the invertebrates they are concerned with transmitting postural information and are present in crustacea, and insects. In insects they are modified cuticular structures in the form of hair, plate, etc. and located on the stretchable area of the cuticle.
ADVERTISEMENTS:
Action of wings in flight is controlled by proprioreceptor cells present at the base of the halteres.
iii. Equilibrium receptor:
Maintenance of body equilibrium is essential for the normal activities in an animal. Equilibrium receptors respond to loss of balance and help the animal to correct it. Lithocysts (Fig. 14.2.), the marginal sense organs, ensure horizontal swimming in cnidarians. Statocysts (Gr. statas = standing + kystis = sac) are ‘organs of balance in arthropods and molluscs.

A statocyst is usually an oval sac, may or may not be in communication with the exterior and lined internally by a sensory membrane bearing pro- arthropods and molluscs. A statocyst is usually an oval sac, may or may not be in communication with the exterior and lined internally by a sensory membrane bearing processes or hairs.
ADVERTISEMENTS:
A single calcareous particle or statolith or a number of small particles, statoconia (Fig. 14.3) are present at the centre.

In some cases, sand particles (Fig. 14.4) occupy the place of statolith. With the change in the body position, the particles exert pressure on certain sensory cells and this signals the animal to correct its position with respect to the gravity. A pair of halteres, the modified hind wings in flies and mosquitoes take care of maintaining stability by controlling lift, rotation and landing in their flight.

iv. Phonoreceptors:
Phonoreceptors or sense of hearing can detect pressure waves caused by mechanical disturbances at some distance from the body.
Three types of phonoreceptors have been identified in arthropods:
1. Sensillae hairs:
ADVERTISEMENTS:
The hairs can perceive low frequency vibrations. They react with the sound waves and are located on the body surface. Frequency of sound waves below 20 hertz is called vibration.
2. Chordotonal organs:
They differ in structures in different arthropods. The type known as tymponal organ, the most complex and efficient organ of sound perception is found in moths. The sensory neurons attached to the apical bodies, the scolopodia are bipolar.
3. Lyriform organs:
It consists of parallel slits of varying length in the cuticle, resembling the strings of a lyre. Nerve endings of sensory neurons are present in the cuticle covering it. Hearing, in true sense of the term, is absent in invertebrates. They can only perceive the disturbances in the medium — air, water or solids.
2. Chemoreceptors:
Receptors for smell and taste are chemo- receptors. These sense organs are capable to differentiate between chemicals and thereby help in controlling the activities of animals in relation to its surroundings, in the selection of food, in social behaviour, in defence from predators, in selection of mate and others.
The mouth parts and terminal segment of butterflies and moths bear hairs and pegs. Receptors for taste and touch are present in these structures. The taste receptors in flies are specific for sugar, water and salts. Not much is known about sense of smell in the invertebrates. The antennae and palpi in insects bear receptors for smell.
Olfactory receptors in moths and ants are quite powerful. The pulmonate molluscs move towards the food, even after removal of both the eyes by surgery. This suggests some sort of sense of smell are present in them. Osphradium in molluscs can test the quality of water and it is considered as a chemoreceptor.
3. Photoreceptors:
Sense organs capable to respond to light stimulus are photoreceptors. Photoreceptors in different degrees of perfection are present in the majority of the animals. Even the protozoon, in which such an organelle is lacking, respond to light and move away from bright light. In Euglena, the photoreceptor, stigma, simply a mass of granules (Fig. 14.5) is present near the base of the flagellum.

In the metazoan, portions of the skin are specialised to form eyes. Clusters of simple eyes or ocelli in the jelly fishes help them to orient to light. Isolated light sensitive cells are scattered over the body surface in earthworm. Bowl-shaped, black pigmented ocelli containing clusters of photosensitive cells are present in planaria (Fig. 14.6).

The pigment allows light to reach the receptors only from above and slightly to the front, but image formation does not occur. Eye spots are present in leech. The eyes of nereis (Fig. 14.7) are more specialised them those of planaria and leech.

A lens to concentrate light and a great increase in the number of photoreceptor cells led to the evolution of image forming eyes.
Two types of eyes—ocelli or simple eyes and compound eyes, are found in arthropods.
Simple Eye:
The ocelli type eyes are constituted by a single ommatidium, as in arachnids (Fig. 14.8) and compound eyes in crustacea and insects. The eyes of cephalopods are camera type like those of the vertebrates.

Compound eye:
A compound eye is formed of a number of visual elements, ommatidia and visions are of two types, mosaic and superimposed.
4. Thermoreceptors:
All animals are sensitive to variations in temperature. No specialised thermo receptors are on record in invertebrates. The thermo receptor cells are usually scattered all over the body surface and may be concentrated in certain regions. e.g., in the antennae of some insects, particularly warm blood sucking forms.
Receptors in Vertebrates:
1. Mechanoreceptors:
Five types of mechanoreceptors are found in vertebrates:
i. Tactile Receptors:
The whole body surface forms an organ of touch. Special tactile receptors are widely distributed on the body surface. In human, extreme concentration of touch receptors are in the fingertips and lips.
Different types of touch receptors (Fig. 14.9) are:
a. End bulbs (Fig. 14.9A) consist of ovoidal groups of sensory cells innervated by a special nerve.
b. Touch cells (Fig. 14.9B) formed by termination of a sensory nerve in the dermis.
c. Touch corpuscles (Fig. 14.9C) ramified sensory nerve in a mass of connective tissue, the terminal branches ending in touch cells.
d. Pecinian corpuscle (Fig. 14.9D) the naked axon is surrounded by a. complex, laminated sheath containing fluid.
ii. Proprio Receptors:
These are internal receptor organs present in skeletal muscles, tendons and joints. They are capable of registering muscle tension or stretch and ensure harmonic contraction of all the muscles involved in a single movement. Proprioreceptors help us to work in dark. Non-functioning of proprioreceptors causes numbness of an organ.
iii. Equilibrium Receptors:
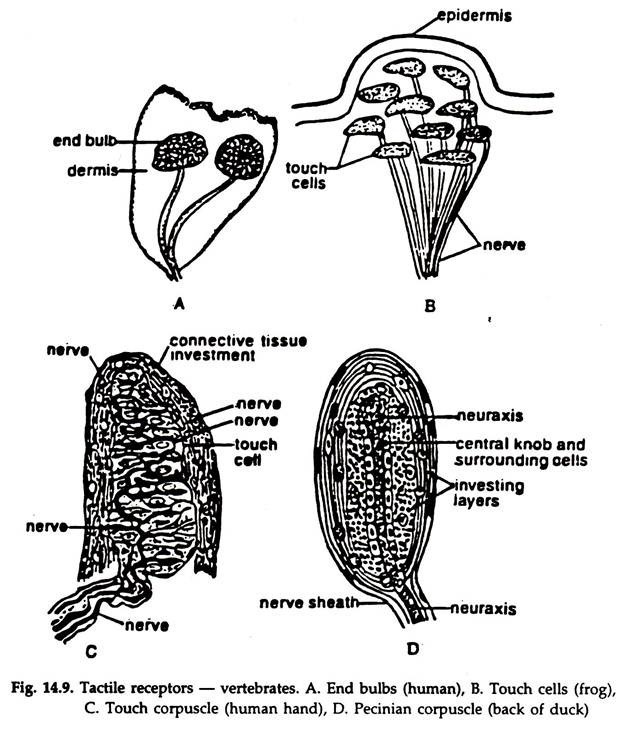
The ability to differentiate the orientation of the body in relation to its surroundings and to maintain an equilibrium is present in all vertebrates. The internal ears or membranous labyrinths control these functions. The membranous labyrinth is enclosed in a cavity, the bony labyrinth and bathed in a fluid, the perilymph, which is protective and acts as a buffer.
The membranous labyrinth (Figs. 14.10, 14.11) consists of three semicircular canals (except agnatha) and two small, hollow sacs, the utricle and saccule. The semicircular canals are connected at both ends with the utricle and arranged in a fashion that each is at right angles to the other two and each canal bears a small ampulla at the lower end.
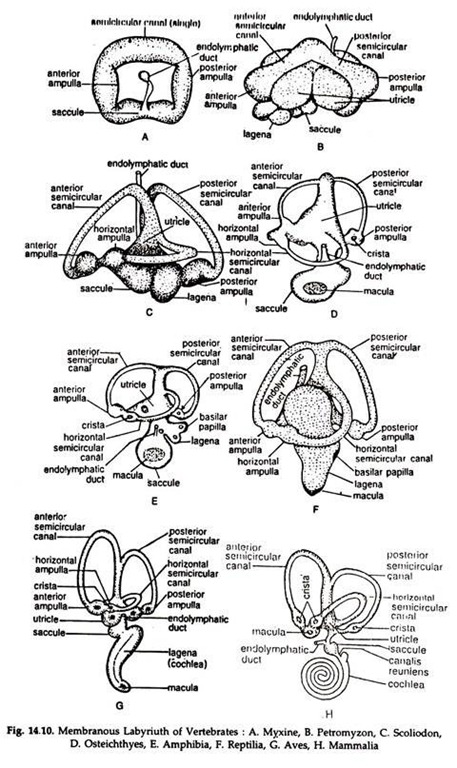
The anterior and posterior canals are vertical in position while the external one is in horizontal plane. Patches of elongated sensory cells produced into hair-like processes are present in the ampullae, utricle and saccule.
They are known as cristae acoustica in utricle and saccule and cristae ampularis in the ampulla. The sensory cells are innervated by the branches of the auditory nerve. The utricle and saccule contain small, calcareous otoliths.
The membranous labyrinth is filled with a fluid, the endolymph. The utricle and saccule are concerned with the position of the head. The flow, of endolymph in the semicircular canals stimulates the sensory cells in the ampulla.
Maintenance of equilibrium:
Change of head position (static equilibrium) affects the gravitational pull on the otoliths and hence the pressure on the sensory hair cells in the utricle and saccule in relation to gravity.
Otoliths are pushed back upon certain hair cells of the utricle and saccule during rapid forward movement (linear accelaration).
The hair cells in the ampulla move faster than the endolymph due to the inertia of the lattter in sudden turns of the head (angular accelaration) and the hair cells are stimulated due to differential movement.
The impulses are transmitted to the brain and the person becomes conscious of the body position.
In addition to the membranous labyrinths, eyes and proprioreceptors help in maintaining equilibrium.
iv. Phonoreceptors in Animals:
The ear is the organ of hearing. The phonoreceptor or the structure concerned with hearing is inseparable from the inner ear or membranous labyrinth but the latter does not take part in hearing. A complete history of transformation of simple ear of fishes to the complex ear of mammals is available.
Water is a dense medium and the sound waves moving in water have a greater impact. The auditory organ in fishes is a simple, blind diverticulum, the lagena at the posterior end of the saccule (Fig. 14.10). With the change of surroundings from water to air on land, the organs of hearing underwent profound changes.
The air being a lighter medium the need to amplify the force of sound waves was a must and this led to the development of new structures. The first new acquisition in amphibia was the ear drum or tympanum on the body surface, which enclosed a space, the middle ear, external to the internal ear.
An oval window, the fenestra ovalis communicates the middle ear with the inner ear. To transfer sound waves from the tympanum to the inner ear, a new structure, as columella anris in amphibians, reptiles and birds and as stapes in mammals was formed from the hyoid arch, which was rendered functionless with the origin of autostyly.
Further, in mammals, incus and malleus in the middle ear developed from the quadrate and articular, respectively.
The ear in mammals though basically similar to that of other tetrapod’s, is more elaborate. The tympanum shifted from surface to inside and the passage from it to the exterior, the external ear appeared.
The cochlea, the part of the membranous labyrinth concerned with hearing is an outgrowth from the saccule. The capability to detect sound waves at different frequencies varies in different animals. It is 20 to 20,000 hertz in man, over 100,000 in bats.
The mammalian ear:
The mammalion ear consists of three distinct parts — external ear, middle ear or tympanic cavity and internal ear (Fig. 14.11).
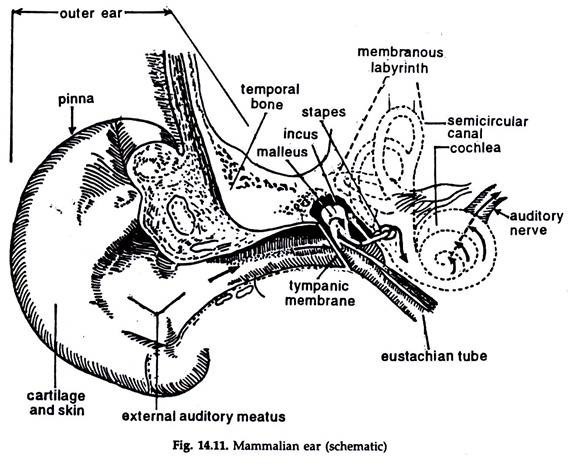
The external ear:
It consists of two components, the skin covered, cartilagenous pinna or auricle and the slightly S-shaped auditory canal or external auditory meatus extending from auricle to tympanum. The tympanum completely separates the external auditory meatus from the middle ear.
The middle ear or tympanic cavity:
The cavity is lined with either a simple squamous or a cubical epithelium. It is filled with air coming from the nasopharynx through the auditory or eustachian tube lined with a ciliated epithelium. Atmospheric air pressure on both the sides of the tympanum allows it to vibrate in response to sound waves.
The medial wall of the tympanic cavity bears two openings, the fenestra ovalis (already referred to) or oval window and fenestra cochlea (round window). The stapes and a fine sheet of fibrous tissue occlude the fenestra ovalis and the fenestra cochlea, respectively.
Auditory ossicles:
Three small bones from outer to inside— malleus, incus and stapes, the last one fitting into the fenestra ovalis constitute auditory ossicles.
The internal ear:
Three structures — the vestibule, the membranous labyrinth and cochlea lodged in a cavity, the bony labyrinth constitute internal ear.
a. The expanded part adjacent to the middle ear and containing the oval and round windows constitute the vestibule.
b. The membranous labyrinth consists of three semicircular canals, utricle and saccule.
c. The cochlea (Gr. kochlias = snail), an outgrowth of the saccule is a spirally coiled tube of two and half turns.
In a cross section, the cochlea is triangle-shaped. The base of the triangle or the basilar membrane is lined by neuroepithelial cells and nerve fibres. The neuroepithelial cells are arranged side by side and a majority of them are long and narrow (Fig. 14.12). These cells and their nerve fibres constitute the organ of corti, the hearing organ proper. The organ of corti is innervated by the auditory nerve.
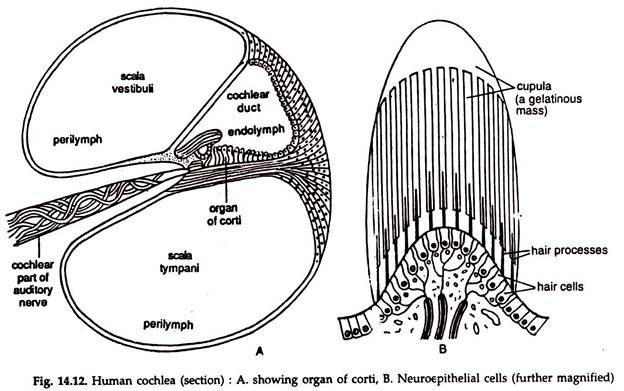
Mechanism of hearing:
Sound waves travel at about 1,088 feet (332 metres) per second. The sound waves are concentrated and directed by the auricle through the external auditory meatus. The tympanic membrane vibrates in response to the pressure of the sound waves.
The vibration of the tympanum is transmitted to the oval window through the middle ear by the ossicles. To and fro movements of the stapes create pressure waves in the perilymph, which, in turn, causes wave motion in the endolymph.
The neuroepithelial cells of the organ of corti are stimulated by the motion and the nerve impulses are send to the hearing area of the cerebrum in the brain through the auditory nerve. Vibrations of the membrane of the round window neutralize the wave motion in the endolymph.
v. Rheoreceptors:
Rheoreceptors or detectors of low vibrations in water, are present in fishes and aquatic amphibians. These are also known as lateral line sense organs. Two lateral line sense organs, each with a series of lateral line organs or neuromasts (Fig. 14.13) constitute rheoreceptors.
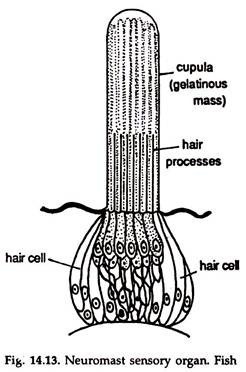
These are located within lateral line canals divisible into (a) a trunk and a tail canal and (b) a head or cephalic canal. The lateral line canals he in the dermis or deeper and covered by the skin bearing numerous pores.
A neuromast is a group of sensory cells with supporting cells. Each sensory cell has a -hair projecting outside through the pore. A mass of gelatinous material, the cupula, secreted by the neuromast cells encloses the tips of the cells.
The sense organs are innervated by the facial nerve and the lateralis branch of the vagus. The organs detect low vibrations in water and enable the fish to move correctly in turbid water and also in darkness.
2. Chemoreceptors:
The sense of smell (olfactory) and the sense of taste (gustatory) constitute chemoreceptors. The olfactory receptors can detect objects from a distance while the gustatory receptors are active only in contact with the substance.
Olfactory receptors:
The olfactory receptors are located in the posterior part of the nasal cavities. The receptors or the olfactory epithelium (Fig. 14.14) is made of olfactory cells, sensitive to smell. One end of the cell responds to the chemical’ stimuli while the other end is continuous with a neuron. The nerve fibres join to form the olfactory nerve, which passes directly to the olfactory lobe of the brain.
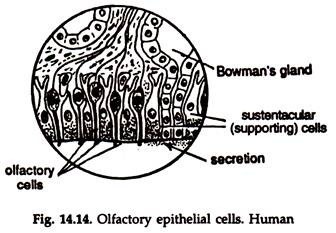
It is believed that substances containing volatile particles when dissolved in the fluid of the nasal cavity, releases the volatile particles, which stimulate the receptor cells. This initiates impulses, which are transmitted to the site of smell in the brain.
In many air breathing vertebrates (amphibians, reptiles, mammals) an offshoot of the olfactory organ separated from it, forms a distinct sac lined by sensory epithelium and innervated by the olfactory and trigeminal nerve. It opens into the mouth and known as Jacobson’s organ (Fig. 37.23).
Gustatory receptors:
The sense organs for taste are bud-like structures, known as taste buds (Fig. 14.15).
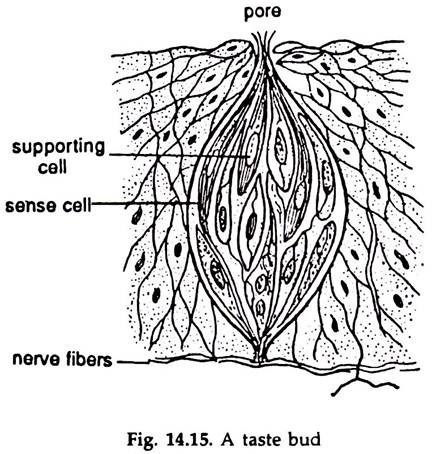
In fishes the taste buds are numerous and located in the mouth cavity, pharynx, skin of the head, tentacles, branchial chambers and, in some cases, scattered on whole surface of the body. In mammals, the buds are located on the papillae in the tongue and soft palate.
The taste buds are derived from the epithelium and oval in shape. Each bud contains two types of narrow cells, the taste cells and supporting cells. The free ends of the taste cells bear microvilli. The bud is in communication with the surroundings through a minute pore and many microvilli project into the pore. The taste bud is innervated by glossopharyngeal nerve.
A taste cell may be connected with only one neuron or more than one. The taste cells are stimulated by substances in solution. Sweet, sour, salt and bitter are four basic tastes. Some buds are specific while others may be stimulated by more than one tastes. This makes the detection and processing of information in the tongue very complex. But the messages finally transmitted to the brain are different for different tastes.
3. Photoreceptors:
Two types of photoreceptor organs or eyes are found in vertebrates. Present day vertebrates possess a pair of image forming, simple, lateral eyes. The ancestral forms had another type of eye, the median pineal eye, which still persists in cyclostomes and some reptiles.
Pineal eye:
The pineal eye of cyclostomes can possibly distinguish light and darkness. It is a round capsule with a lens formed of elongated cells at the anterior wall and the posterior wall resembles a retina, made of an inner layer of rod-like visual elements and an outer layer of nerve fibres (Fig. 32.12B). In reptiles (Fig. 14.16), the nerve degenerates in the embryo and the eye lost its function.
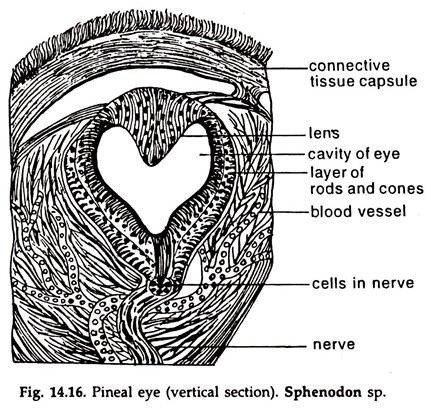
Image forming eye:
The vertebrate eyes are camera-type eyes. Since the vertebrates live in different media, their eyes are also adapted to see under water, in bright light, in dim light and in darkness (in relation to human vision). Still the basic features and functions of different components are similar in all groups of vertebrates.
Mammalian eye:
Eyes or organs of vision are two in number in a mammal, like that of any other vertebrate. The eye lies safely in a bony orbit of the skull, excepting a small exposure in front bounded by the eyelids.
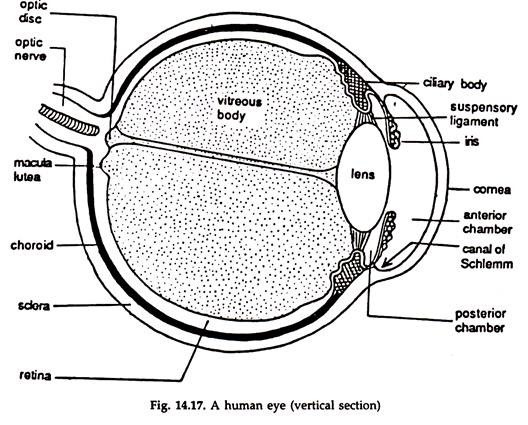
It is a hollow ball-like structure with a lens in front and a sensitive screen behind and largely corresponds to an ordinary photographic camera (Fig. 14.17). The eye is moved by six muscles— superior oblique, inferior oblique, superior rectus, inferior rectus, anterior rectus and posterior rectus.
Structure:
The wall of the eye-ball has three concentric layers—outer tunica fibrosa, middle tunica vasculosa and inner retina.
Tunica fibrosa:
The outermost layer of the eye-ball, tunica fibrosa, skeletal in function, and maintains the rigidity of the eye-ball. It consists of sclera and cornea. The sclera is opaque, and is supplied by optic nerve and blood vessels. In the front, the sclera continues as a clear, transparent membrane, the cornea. A thin mucous membrane, in continuation with the eyelids, is present in front of the cornea. This is the conjunctiva.
Tunica vasculosa:
The layer beneath the sclera is called tunica vasculosa. It is composed of choroid, ciliary body and iris, rich in blood vessels and pigment cells.
Choroid:
The inner posterior five-sixths of the sclera is lined by choroid. The choroid is pigmented and richly supplied with blood.
Ciliary body:
The anterior continuation of the choroid is made of non-striated muscle fibres, the ciliary muscle and known as ciliary body. It is attached to the suspensory ligament, the other end of which is continuous with the capsule enclosing the lens.
Iris:
The iris is a circular body behind the cornea, in front of the lens and with an aperture at the centre, the pupil. The iris is composed of two layers of muscle fibres, one circular and the other radiating and contain pigment cells.
Contraction of the circular and radiating muscle fibres constrict and dilate the pupil. The space between the cornea and the lens is filled with a watery fluid, the aqueous humor. The iris incompletely divides the space into a large anterior and a very narrow posterior chamber.
Lens:
The lens lies just behind the pupil. It is a transparent, circular, biconvex, highly elastic body. The lens is enclosed in a transparent capsule and suspended from the ciliary body by suspensory ligament. Contraction of the ciliary muscles release the pull on the lens and it thickens and moves forward.
Retina:
It is the innermost layer of the eye wall. The retina lines about posterior three-fourths of the eye-ball. It is thickest at the posterior end, becomes thinner anteriorly and terminates just behind the level of the ciliary body.
The retina is composed of a layer of pigmented epithelium constituted by receptor cells, the rods and cones (Fig. 14.18), against the choroid and several layers of sensory and connector neurons and their axons on the inner side.
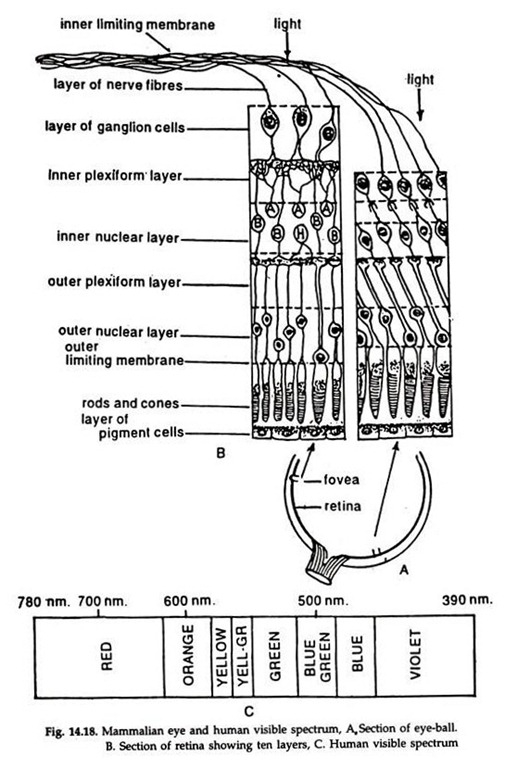
Due to the back position of the light sensitive cells, the rods and cones, the light rays must pass through several layers of neurons to reach them. The rods are more numerous and sensitive to dim light and colours while the cones are less numerous and respond to bright light. The retina acts as a sensitive screen on which the images of objects are focused. The rods and cones receive the visual impulses.
The retinal layer is highly innervated by the optic nerve. The point where the optic nerve enters the retina is known as blind spot. Situated close to the blind spot is the macula lutea (yellow spot).
A small depression, the fovea centralis at the centre of this region is the site of greatest visual activity. Fovea centralis contains only cones. The high activity is due to the presence of maximal number of receptor cells, all cones, in the area.
Vitreous body:
The whole space between the lens and the retina, about four-fifths of the eye-ball, is occupied by a thin jelly, the vitreous humor, enclosed in a delicate, transparent membrane, the hyaloid membrane. A narrow canal, the hyaloid canal, filled with lymph runs forward through the vitreous body from the entrance of the optic nerve to the posterior surface of the lens.
Functions:
Light rays enter the eye-ball through the transparent cornea and are focused by the lens on the retina. An inverted image of the object is formed on the retina and this is conveyed to the visual centres of the optic lobes in the brain by the optic nerves. The inverted image is corrected and a direct image is formed. The visible spectrum of man ranges from violet (390 nm) to red (780 nm).
Accommodation:
Normally, the eye is focused for distant vision. It can be adjusted for near vision by increasing the convexity of the lens. Due to the tendency of the eye-ball to remain globular normally, there is a pull on the lens which keeps it flat.
Contraction of the ciliary muscles releases this pull on the lens and the elastic lens becomes more convex and moves forward. The divergent rays from the near object are thus focussed to form a clear image on the retina.
4. Thermoreceptors:
Cells sensitive to temperature variations are present in all vertebrates. In most cases, they are scattered all over the body. Fishes are highly sensitive to temperature variations and can detect variation as low as 0.1°C.
This enables the fishes to know that they are entering a different body of water and to regulate their migration. In snakes, the thermo receptors are usually located in pits on the sides of the head. This helps them to locate the presence of warm-blooded prey.
5. Electroreceptors:
Electroreceptors are sensory cells which can detect changes in the electric field of the surroundings. The fishes, electric ray or electric eel can generate electric pulses exceeding 500 volts. The majority of the fishes having electric organs, however, give off very weak pulses.
These are believed to be a part of the electric guidance system. It is presumed that with their specific electric organs the fishes create a pulsating electric field around them and any object in the surroundings with a different electrical conductivity than that of the water will disturb the field. With their special receptor cells these fishes can detect the disturbance in the electric field and alter their course in water.