The following points highlight the top two types of respiration in mollusca. The types are: 1. Aquatic Respiration 2. Terrestrial Respiration.
Type # 1. Aquatic Respiration:
In this, O2 dissolved in water is used. The surface of the respiratory structure is greatly increased to allow rapid absorption of O2.
The aquatic respiratory structures may be placed under two heads—primary and secondary:
Primary respiratory structures:
ADVERTISEMENTS:
The primary respiratory structures are ctenidia or gills. The gills are located in the mantle cavity, attached to the body by membranes.
Structure of ctenidia:
1. Ctenidia are paired, symmetrical, ciliated and two rows of flattened gill filaments, arranged one on either side of a long, flattened axis, traversed by afferent and efferent vessels through which haemolymph flows.
2. Narrow spaces are left between the gill filaments to permit free water flow between them but close enough so that cilia on adjacent filaments may operate together in generating water current.
ADVERTISEMENTS:
3. Haemolymph circulates through the gill filaments.
4. Skeletal rods support the gill filaments.
5. Cilia generate inhalant water current drawing water below the ctenidia and exhalant current expelling water out from above them.
Types of ctenidia:
ADVERTISEMENTS:
Based on topography, the ctenidia in molluscs are placed in following categories:
1. Holobranchiate:
The ctenidia extend all over the body. The number varies from 14-80 pairs. Example: Polyplacophora.
2. Merobranchiate:
The ctenidia are restricted to a particular area of the body.
Depending on the arrangement of leaflets the gills have been subdivided into four types:
A. Plicate:
Simple, flat, transversely folded, projecting integumentary laminae constitute the gill. In Neomania a tuft of filaments arise from the cloacal wall.
B. Monopectinate:
ADVERTISEMENTS:
Flattened gill filaments are arranged in a single row (Fig. 26.16). Examples: Pila, Triton, etc.
C. Bipectinate:
Flattened gill filaments are arranged in two rows.
They are of two types:
a. Unequal:
Both rows of filaments are present, the right one being smaller in size. Example: Fissurella.
b. Equal:
Both the rows of filaments are of same size.
This type is present in Bivalvia, but they are variously modified:
i. The leaflets are short and flat.
Example: Nucula.
ii. The leaflets are filamentous and long.
They may be free (Area) or joined by ciliary connectives (Mylilus).
iii. The ciliary junctions are replaced by membrane.
Example: unio (Figs. 26.2, 26.5).
iv. The gill is degenerated and represented as transverse partitions.
Example: Porontya.
D. Feathered:
Assumes the shape of a feather. (Figs. 26.35, 26.38) Examples: Cephalopods.
Ctenidia in different groups of molluscs:
1. Monoplacophora:
The pallial groove contains five to six pairs of unipectinate gills.
Example: Neopalina.
2. Polyplacophora:
Bipectinate ctenidia located in the mantle groove:
(a) Number of ctenidia varies from 14 (Lepidopleura) to 80 (Acanthopleura).
(b) Gill rows may be holobranchiate with two exceptions bearing merobranchial type. Examples: Chiton laeves, Chiton pall as ii.
3. Aplachophora:
The gills are reduced to a paired, feather-shaped structure situated near the cloacal cavity, one on each side. The gills are merobranchiate. Example: Chaetoderma.
4. Gastropoda:
The gills vary widely in number and position in this group:
A. Prosobranchia:
Ctenidia lie in front of the heart.
a. Diatocardia:
The arrangement is most primitive. The gills are two, long, feathered, on each side and lie symmetrically to the middle line. Example: Fissurella.
b. Monotocardia:
The arrangement of the gills is remarkably uniform. A single gill, feathered on one side and united to the mantle along its whole length. Example: Triton.
B. Opisthobranches:
The gills are partially enclosed in the mantle cavity (Tectibranchiata). The true ctenidium, when present, is little developed and located on the right side of the body.
Example: Aplysia.
5. Bivalvia:
The gills are bipectinate, equal on either side, usually very large, having assumed food collecting function in most species in addition to gas exchange.
A. Protobranchiata:
Gills are smallest and lie behind the foot at the back of the mantle cavity. In Nucula the gill filaments are triangular.
B. Filibranchiata:
Each gill separately forms a ‘W’ in section-.with long, narrow limbs. The gill axis lies at the middle angle of the W. Examples: Area, Mytilus, Anomia.
C. Pseudolamellibranchiata:
The reflected dorsal tips of the gill filaments have coalesced laterally with the mantle and mesially with the base of the foot. The gill has a greater cohesion than that in the filibranchs. The gills are more complex, with the surface plicate or thrown into folds and grooves. Examples: Ostreidae, Pectinidae, Pleriacea.
D. Eulamellibranchiata:
The adjacent filaments of gills are united by vascular cross connections, leaving narrow openings, the Ostia between them. The two lamellae of each demi-branch are attached back to back in the same way. Examples: Cardiacea, Myacea.
E. Septibranchiata:
The ctenidia do not exist as such, being converted into a horizontal muscular septum running from the base of the foot to the mantle, and extending right back to the siphons. Example: Poromya.
6. Cephalopoda:
The gills are large, paired, bipectinate and one suspended on either side of the rectum by their afferent edges, not by efferent, as in gastropods, and the water current is driven from afferent to efferent side.
A. The gill filaments are firm and fleshy, non-ciliated, and thrown into primary and secondary folds to increase respiratory surface. In Nautilus, the gills are two pairs.
B. The haemolymph flows through each gill is assisted by a pulsatile accessory branchial heart, located at the base of the gill in the course of the afferent ctenidial vessel and placed in an annexe of the pericardium.
C. Pallial contractions drive water between the gill filaments at great pressure.
Evolution of ctenidia:
The most primitive ctenidia are found in zeugobranchiate prosobranchs, which are simple outgrowths of the body. In bivalves;
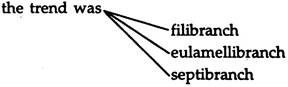
with an increase in length of the ctenidium. In this group three kinds of new tissue connections evolved.
1. Interfilamentar junction between adjacent filaments.
2. Inter-lamellar junction between two lamellae.
3. Connections between the tips of the filaments and mantle or foot.
In bivalves, the branchial apparatus in addition to respiratory function acts as a food collecting organ. Large, non-food particles are trapped and strained out by the mucus secreted by hypobranchial gland. Osphradium tests water quality.
Adaptive or secondary gills and integument:
The gills develop from aberrant sites:
1. Anal gills:
Delicate leaflets form a rosette around the anus. Example: Doris. In Chactoderma a pair of symmetrical lateral gills are present on each side of cloaca.
2. Cerata or dorsal appendages:
In many opisthobranches the dorsum bears highly vascular appendages called cerata. They may be simple and club-shaped (Aeolis), dendritic (Dendronotus) or multi-lobed resembling a bunch of grapes (Dotochica).
3. Pleural gills:
In Pleurophyllida lateral rows of branchial leaflets are situated beneath the mantle.
4. Pallial gills:
In certain basommatophore pulmonate secondary external gills develop by the enlargement of the pallial lobe, just outside the pneumostome. They, however, lack cilia. Examples: Planorbidae, Ancylidae.
5. Integument:
Specialised respiratory structures are lacking in some Scaphopoda. Respiration is carried by the internal surface of the mantle, particularly the anteroventral side in Dentalium, Antalis. In nudibranchs (Gastropoda) the entire dorsum of the body acts as the site of gas exchange. Integumentary gas exchange occurs in parasitic Entoconcha, Conia, Limpontia sp. etc.
Type # 2. Terrestrial Respiration:
Amphibious adaptation and adaptation to terrestrial life have introduced remarkable modification in the respiratory structures of molluscs.
The structures associated with terrestrial respiration are:
1. Nuchal lobe:
The left nuchal lobe is better developed and forms a long respiratory siphon.
Example: Monotocardia.
2. Pulmonary sac:
In some amphibious prosobranchs—Pila. (Fig. 26.17), Ampullarius, Siphonaria, etc. the pallial cavity is incompletely partitioned by a fleshy fold, the epitaenia, into a right branchial chamber and a left pulmonary chamber. The highly vascularized roof of the pulmonary chamber forms a pulmonary sac, the aerial respiratory structure, with a small aperture opening in the pulmonary chambers.
Air enters the pulmonary chamber through an opening at the tip of the left siphon. Aerial respiration takes place during aestivation and when there occurs a depletion of O2 concentration in water. The pulmonary epithelium comprises cuboidal or squamous cells bearing short microvilli and many mucous cells.
3. Lung:
In pulmonate — both stylommatophores and basommatophores the — respiratory structure is a true lung of independent origin, not vascularized mantle, which occupies the major part of the roof of the pallial cavity.
In some it extends to the walls and floor. In slugs Athoracophoridae the lung is nonvascular and its wall gives off a number of delicate branched tubules, named tracheae (Hiscock, 1972), which function as respiratory structures.
The pallial cavity opens to the exterior by a large, oval aperture, the pneumostome, the opening and closure of which are controlled by contraction and relaxation of muscles. Large pulmonary veins with extensive ramification drain oxygenated blood from lung to auricle.
Respiratory pigment:
The respiratory pigment is an extra corpuscular haemocyanin. It is more concentrated in gastropods and cephalopods. In Area, and Solen the pigment is haemoglobin and present in some special corpuscles in the haemolymph.
Mechanism of Respiration and Oxygen uptake
A. Aquatic respiration:
Water is driven over the ctenidial surface by ciliary beating or by muscular pumping.
a. Cephalopoda have branchial hearts at the gill bases, the pulsation of which drives haemolymph through afferent vessels.
b. Lack of such additional pump and less efficient funnel contraction are compensated with the development of two pairs of ctenidia in Nautilus.
c. Oxygen utilisation from pallial water is high in gastropods and cephalopods. It is 56% in Haliotis, 79% in Triton and 63% in Octopus, as compared with 5-9% in sedentary lamellibranch’s.
B. Terrestrial respiration:
Contraction and relaxation of muscles in pulmonates cause in and outflow of air into and from the pallial cavity.
a. In a relaxed state of muscles the floor of the pallial cavity (diaphragm) arches upward.
b. During inspiration the pneumostome opens, muscular contraction flattens the floor causing fall of pressure in the pallial cavity and air rushes in. This is followed by closure of the pneumostome.
c. A slight delay in the opening of the pneumostome facilitates O2 uptake by haemolymph.
d. Respiratory movements increase with lowering of the O2 concentration and at higher temperature. Such movements have been reported from Arion, a slug with a large pneumostome (Runhon & Hunter, 1970).
e. In prosbranchs alternate dilatation and contraction of the mantle and pulmonary chamber wall help in and outflow of air in the pulmonary chamber through left siphon, which enlarges to reach above water level when the snail remains submerged.
Molluscs are sluggish creatures except cephalopods. The lower respiratory activity in the majority, however, is sufficient to meet with the need of energy of the animals. In cephalopods, a rapid flow of haemolymph through the gills is effected with the development of accessory branchial hearts, helping higher uptake of oxygen by the gills.
The presence of diverse but well-organised respiratory structures—gills and lung—and certain other features led some authors to believe that molluscs are di-phyletic in origin.