In this article we will discuss about osmoregulation in various kinds of fishes.
According to habitat, fishes can be distinguished as (i) Marine, and (ii) Fresh water.
(i) The marine fishes fall into two distinct groups, (a) Those whose osmotic concentration is the same as or slightly above sea water, e.g., hagfish, elasmobranchs, Latimeria etc. This group has no major problem of water balance, because its inside and outside concentrations are equal, there is no osmotic water flow, (b) Those whose osmotic concentrations are about one third of that of sea water, e.g., lampreys, teleosts, etc. These are hyposmotic animals. They live in constant danger of losing water to the osmotically more concentrated medium.
(ii) The fresh water fishes, on the other hand, have internal concentrations greater than that of their external medium. Thus, they are hyperosmotic to the medium. Therefore, the osmotic problems and the means to solve them differ drastically among fishes of different habitats. A list of some fishes with habitat and osmotic characteristics is given in Table 8.9.

Osmoregulation in Cyclostomes (Lampreys and Hagfishes):
The cyclostomes have two groups:
(i) Lampreys, which are anadromous, i.e., live both in sea and in fresh water,
(ii) Hag- fishes are strictly marine and stenohaline. The Lampreys, weather fresh-water or marine, have osmotic concentrations about one-quarter to one-third the concentration of sea-water. Their main problem is similar to that of teleost fish.
These problems are discussed later. The hagfishes are the only true vertebrates whose body fluids have salt concentrations similar to that of sea-water. In fact, the normal Na+ concentration in hagfish blood exceeds that in their surroundings (Table 8.9). Therefore, they have pronounced ionic regulation as an isosmotic animal.
ADVERTISEMENTS:
Osmoregulation in Marine Elasmobranchs:
The common examples of marine elasmobranchs are sharks and rays. The salt concentration in their body fluid is roughly one- third the level of the sea-water (Table 8.9), but they still maintain osmotic equilibrium. This is achieved by adding to the body fluids large amount of organic compounds primarily urea. Addition of different organic compounds in the body fluid/blood makes the osmotic concentration equal or slightly above the sea-water (Table 8.9).
In elasmobranchs urea is a normal component of all body fluids; this is abnormal for other vertebrates. In marine elasmobranchs, the tissue cannot function normally in the absence of such a high urea concentration. Urea is the end product of protein metabolism in vertebrates. Generally it is excreted through kidney, but the shark kidney actively reabsorbs this.
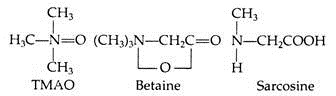
The use of urea for maintaining osmotic equilibrium helps these animals to keep salt concentration much lower than those of sea-water (Table 8.9). But urea can pose problems in the body functioning. It is known that urea destabilizes many proteins, especially enzymes. This problem is solved in elasmobranch by the presence of another organic substance trim-ethylamine (TMAO). TMAO inhibits the effect of urea on enzymes.
ADVERTISEMENTS:
Some other urea inhibiting compounds are betaine and sarcosine:
Although the eldsmobranchs have solved the osmotic problem of life in the sea by being isosmotic, they are still capable of extensive ionic regulation. It is clear from Table 8.9, that they have sodium concentration much lower than that of sea-water. This means that sodium tends to diffuse from sea-water into the body.
Sodium enters into the body primarily through the thin epithelium of the gill and then through the ingested food. This enhances the sodium level in the body, which must have to be excreted or eliminated from the body.
Part of the sodium excretion is undertaken in the kidney. Major excretion of Na+ is performed by a special rectal gland. This is a small gland that opens via a duct into the posterior part of the intestine, the rectum. The gland secretes a fluid with high sodium and chloride concentrations, which is higher than the sea-water concentration.
The elasmobranch blood is usually slightly more concentrated than sea-water. This higher concentration inside causes a slight osmotic inflow of water via the gills. In this way, the elasmobranchs slowly gain water osmotically, and this water is used for the formation of urine and for the secretion of rectal gland.
The excess osmotic concentration is due to the presence of urea. But retention of urea solves the otherwise difficult osmotic problem of maintaining a low salt concentration while living in sea.
Osmoregulation in Freshwater Elasmobranchs:
Majority of elasmobranchs are marine, few of them are also found in freshwater. Some marine species occasionally enters rivers and lakes for various purposes. Among freshwater elasmobranchs, Carcharhinus leucas of Lake Nicaragua, four species of elasmobranchs of Perak River in Malaysia and Amazon sting ray Potamotrygon, are remarkable.
Their blood concentrations are lower than those of strictly marine forms. The urea concentration is reduced to less than one third of the value of marine sharks.
ADVERTISEMENTS:
The problem of osmotic regulation is reduced due to the low level of solutes in the blood. The osmotic inflow of water is diminished because lower salt concentration is easier to maintain. The reduced osmotic inflow of water gives less water to be eliminated by the kidney.
The urine always contains some solutes; therefore, a low urine flow reduces the urinary salt losses. Smith (1931) argued that, it is difficult to say whether the lowered blood concentration is a primary adjustment or merely a passive result of the increased inflow of water and concomitant urinary losses.
Osmoregulation in Teleosts:
Teleost fishes are living both in marine and freshwater. Both types of fishes maintain their osmotic concentration at about the quarter to one-third the level in sea-water (Table 8.9). There is another type of fish, which roams both in sea water and fresh water.
Therefore, can tolerate a wide range of salinities. These movements are often associated with the life cycle, such as breeding. But this change from one environment to the other requires profound changes in the osmoregulatory processes.
(a) Osmoregulation in marine teleosts:
Marine fishes are hyposmotic with the environment. Their main problem is losing body water to the more concentrated sea-water. The body water comes out through their body surfaces, in particular the large gill surfaces. The gill surface is more permeable to water than general body surface. These fishes compensate their inevitable water loss by drinking sea-water.
Drinking of sea-water may restore the water content of the body, but impose another problem. Along with the sea-water, large amount of salts are also ingested and absorbed through the intestine. So salt concentration of the body increases.
Now the problem becomes elimination of excess salt. To solve the problem, salts must be excreted in a higher concentration than in the water taken in. The teleost kidney cannot serve this purpose, because it cannot produce a urine that is more concentrated than the blood.
Therefore, some other organ must participate in solving this problem of eliminating excess salts. This is done by the gills. So gills have dual function—one, participation in osmoregulation and second, gas exchange. The secretion of salt through gill is an active process, i.e., energy mediated. Because, it takes place from a lower concentration in the blood to a higher concentration in the surrounding medium.
Fig. 8.38 describes the summary of osmoregulation in marine teleosts. Fig. 8.38A shows the movement of water and Fig. 8.38B shows the movement of salts. The excretion of sodium and chloride in the urine is of minor importance because teleost urine is more dilute than the body fluids.
However, the kidney plays a major role in the excretion of divalent ions, magnesium and sulfate. These ions are not eliminated by the gills, which seems to transport only sodium and chloride.

Further studies show that the salt intake is happening not only through drinking sea-water, but also through the general body surface. It is also proved that fish adapted to sea-water are relatively permeable to ions and those adapted to fresh water are relatively impermeable.
The ion transport is carried out, not by the general epithelial cells of the gills, but specifically by some large cells known as chloride cells. These cells are also present in the opercular cover of the fish. These cells actively transport chloride ions.
(b) Osmoregulation in fresh-water teleosts:
The osmotic concentration of the blood of fresh water teleosts in much higher than the surrounding water (~300 mOsm/litre). Therefore, their major problem is the osmotic water inflow (Fig. 8.39A). Water mainly enters through the highly permeable gills. In freshwater teleosts skin is less important in transporting water inside the body, because it is less permeable.

The large volume of water is excreted as urine, which is very dilute and may be produced in quantities up to one-third of the body weight per day. The urine contains 2 to 10 mmol/litre of solutes. So large urine volume also causes a substantial loss of solutes (Fig. 8.39B).
This loss is replaced by the gills, which is also slightly permeable to ions. Some solutes are taken in with the food, but the main intake is by active transport in the gills. It is evident from the studies that skin plays only a minor, if any, role in active absorption.